

1. 서 론
해양 생태계에서 기질은 부착하거나 포복하는 저서생물에게 서식 공간을 제공하는 역할을 한다(Kei 2008). 기질에 정착한 저서생물은 생존에 필요한 먹이 활동과 포식자로부터 자신을 보호하기 위한 과정을 거치며 군집을 형성한다(Kirsten et al. 2010). 또한 가입된 저서생물은 수층에서 부화와 수정 등 변태과정을 거친 유생이 다시 착저나 출아 같은 다양한 과정과 기작을 통해 서식공간을 확대한다. 이때 기질에 가입되는 양상은 유생 공급 시기와 기질의 적합성, 착저 이후 공간 경쟁 등 여러 요인에 의해 영향을 받는다(Connell 1985).
부착성 저서생물은 산업사회에서 선박 또는 해양 시설 등 다양한 해양구조물에 부착함으로써 기능을 방해하거나, 부작용을 유발하여 오손생물(fouling organism)이라고 불리고 있다. 이러한 오손생물의 역할로 인해 부착성 저서생물 연구는 생태적 정보보다는 오히려 대상 기질로부터 부착을 제어하는 기술개발이 중요한 목적이 되었다(Jana et al. 2019; Satheesh and Wesley 2009). 특히 양식 산업에서도 해상에 축양하는 시설에 부착하여, 생산성에 경제적 영향을 미치는 장애요인으로 인식되었고(Dürr and Watson 2010), 가두리 어류 양식에서 그물 망목을 막아 양식생물에게 원활한 해수 순환을 방해하는 요인이 되고 있다(Aarsnes et al. 1990; Loland 1993). 부착성 저서생물의 가입과 서식 요인은 수온, 수심, 조류 등 환경 조건과 더불어 기질의 설치 형태 및 방향, 재질 등이 영향을 준다(Braithwaite and McEvoy 2005; De Nys and Guenther 2009; Fitridge et al. 2012). 그 중 수심은 부착성 저서생물의 분포와 군집구조 형성에 중요한 영향을 미치는데(Witman 1985; Underwood et al. 1991; Gallahar and Kingsford 1992; Witman and Dayton 2001), 수심에 따라 종의 조성이 달라질 수 있고, 변화하는 환경 조건에 대한 내성과 먹이, 산소 등 제한된 자원에서 적응하는 능력을 알아볼 수 있다(Heyns et al, 2016). 또한 부착성 저서생물의 부착시기에 따른 차이는 군집형성 과정 중에 발생하는 먹이 및 공간경쟁 등에 큰 영향을 미칠 수 있다(Underwood and Denley 1984; Underwood and Petraitis 1993; Hutchinson and Williams 2001).
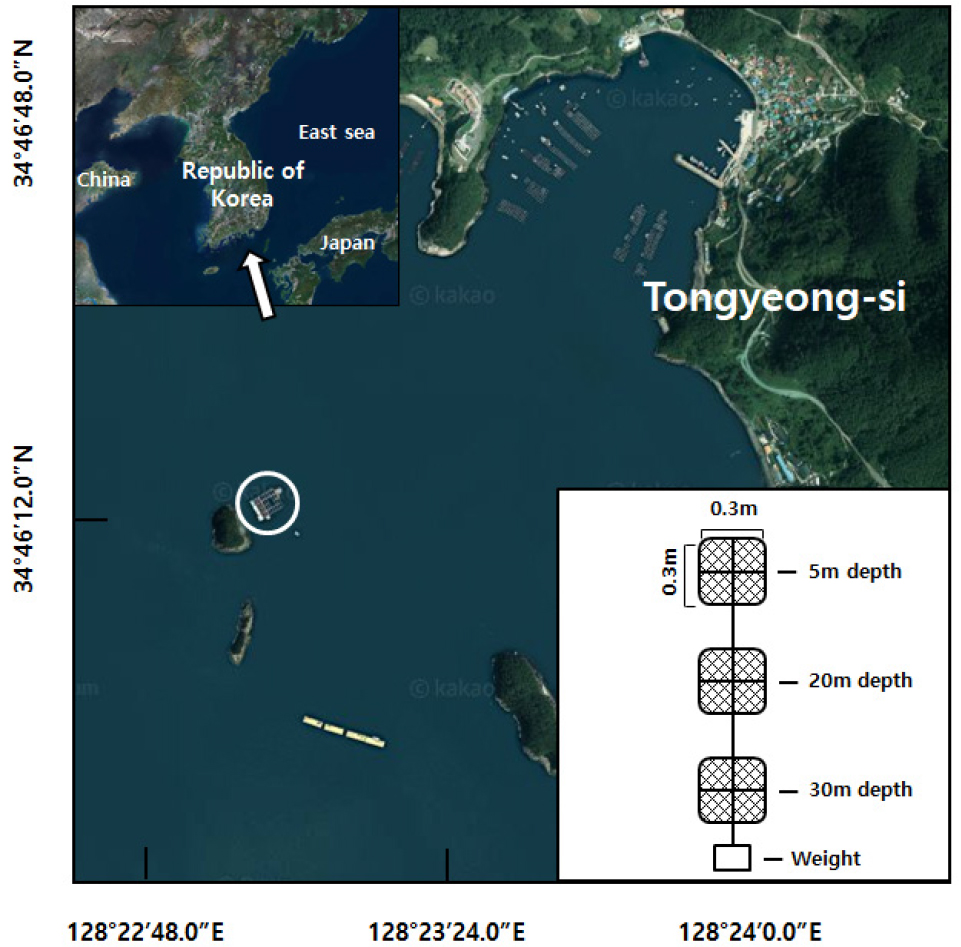
한반도 남해역은 쓰시마난류, 중국대륙 연안수, 한국고유 연안수 등이 계절에 따라 유입되면서, 수온, 염분, 탁도 등에 영향을 미치고 있으며, 복잡한 해안 구조와 조석에 의해 국부적으로 발달한 암반 해안과 갯벌 등 시·공간적으로 다양한 연안 환경 조건이 매우 복잡한 연안생태계 구조를 나타내는 해역이다(양 등 1998; 윤 2017). 통영 주변 해역도 복잡한 해안선과 많은 도서를 포함하고 있으며, 주요 하천은 없고, 외해와 개방된 해역특성을 가지고 있다(윤 등 2010). 지형학적 특성으로 인해 외해로부터 차단되어 물리적 조건으로부터 비교적 안정된 해상 공간에서는 양식 활동이 활발하여, 1970년대부터 공간이 밀집된 정도로 어·패류양식이 성행하고 있다(임 등 2010). 통영 해역의 최근 10년간 연간 평균 수온은 16.6°C 이며 최저 7.9°C(1월), 최고 24.7°C(8월)로 나타났고, 표층의 연간 평균 염분는 32.8 mg/L이며, 계절에 따라 29.8~33.9 mg/L 범위를 나타냈다(국립해양조사원 2019). 조사 지역은 2002년부터 바다목장 사업 수행으로 수산 어류 등 종 보존을 위한 가두리 시설물이 위치한 공간으로, 수심이 낙조시 최대 40 m를 유지하고 있다. 장기간 동안 표층에 설치된 가두리 시설물에는 통영 해안에서 서식하는 부착성 저서생물이 군집을 이루면서 부착생물의 유생공급이 비교적 원활할 수 있는 조건을 나타내고 있다.
대상 지역에서 부착성 저서생물과 관련된 연구는 수산생물의 종묘 확보를 위한 생태 연구(허 등 2008; 조 등 2010)와 주요 부착생물에 대한 가입 양상(박 등 2011; 최 등 2011)에 대한 연구가 진행되었다. 이들 연구는 표층과 수심 2 m 이내 부근에서 진행되었고, 부착정도를 파악하기 위한 기질의 종류나 설치 방향을 고려한 군집구조의 차이 또는 부착 양상을 파악하는데 목적을 두었으며, 수심에 따른 상대적인 가입양상이나 부착시기에 따른 군집조성에 대해 비교한 연구는 진행되지 않았다. 본 연구 목적은 해상 가두리 양식에서 사용하는 그물에서 시기에 따라 수심 별로 부착하거나 가입하는 저서생물의 종조성과 군집구조를 파악하고자 하였다.
2. 재료 및 방법
현장 실험은 통영시 산양면에 위치한 대장두도 주변 어류 가두리 시설물(34°46'11.7"N, 128°22'59.2"E)에서 2019년 3월부터 9월까지 7개월 동안 진행하였다(Fig. 1). 부착 기질은 30 × 30 cm 크기의 알루미늄 재질로 정사각형 틀을 제작한 후에, 가두리 양식에서 일반적으로 사용하는 가두리 어망(망목: 1.5 cm), 폴리프로필렌 소재(PP)를 부착하였다. 제작된 부착판은 한 개의 로프에 5 m, 20 m, 30 m 수심에 계류할 수 있도록 각 1개씩 고정하고, 하단에는 20 kg 무게의 추를 달아서 거치하는 동안 조류에 밀리는 영향을 최소화 하였다. 부착 기질은 조석에도 동일 수심을 유지할 수 있도록 가두리 시설물에 고정 시켰다. 부착 기질은 2개월 간격(3, 5, 7월)으로 각각 투입한 후에 2개월이 경과된 5, 7, 9월에 각각 수거하였다. 수거 과정에서 부착기질에 서식하는 생물의 유출을 최소화 하도록 천천히 인양하였고, 부착기질은 수심에 따라 별도로 분리한 상태에서 각각 10% 포름알데히드 용액을 첨가하여 실험실로 이동하였다. 실험실에서 부착기질은 300 ㎛ 망목의 체로 걸러내어 포복성 생물을 채집하였고, 부착성 저서동물의 경우 어망 중에 25% 면적에 해당하는 15 × 15 cm를 절단한 후에, 어망에 부착된 모든 생물을 탈거하였다. 세척과정에서 수집된 포복성 생물과 부착기질에서 떼어낸 생물은 광학현미경을 이용하여 동정한 후, 종 별로 계수하였고, 습중량을 측정한 뒤 70% 에탄올에 보관하였다. 채집한 면적을 보정하여 개체수 및 생체량 정보를 단위면적(m2)으로 환산하였다. 부착생물 중에 군락을 형성하면서 부착면을 피복하는 태형동물 등은 현미경을 활용하여 군체에서 각 개체를 계수하였다. 생물 부착 실험 장소에서 CTD (YSI- 6000)를 사용하여 월 간격으로 수심에 따른 수온과 염분을 측정하였고, 주변 해역의 연간 수온, 염분 변동은 국립해양조사원 바다누리해양정보 서비스의 관측 자료를 참조하였다. 부착생물의 종조성에 대한 시공간 분석과 다양도(Shannon and Weaver 1963), 풍부도(Margalef 1958) 및 균등도(Pielou 1975)를 산정하였으며, 우점종은 서식밀도와 생체량 자료를 근거로 출현 빈도에 비중을 두는 방식인 Le Bris index (Le Bris 1988)를 적용하였다. 군집분석은 다차원척도법(Multi Dimensional Scaling)을 적용하였고, 각 군집에서 주요종의 역할을 해석하는 방법으로 SIMPER 분석(PRIMER ver 6.0)을 실시하였다.
조사기간을 3월에서 9월로 설정한 이유는 수온이 따뜻해지며 생물들의 대사 및 생식 활동이 왕성하고 부착생물들의 가입이 활발히 일어나는 시점이기 때문이고 거치기간을 2개월로 지정한 이유는 부착생물이 가입되고 성체로 발달되어 군집을 형성해 육안으로 확인하기까지 시간이 최소 2개월 이상 소요되는 것을 감안하였다.
3. 결 과
수층 특성
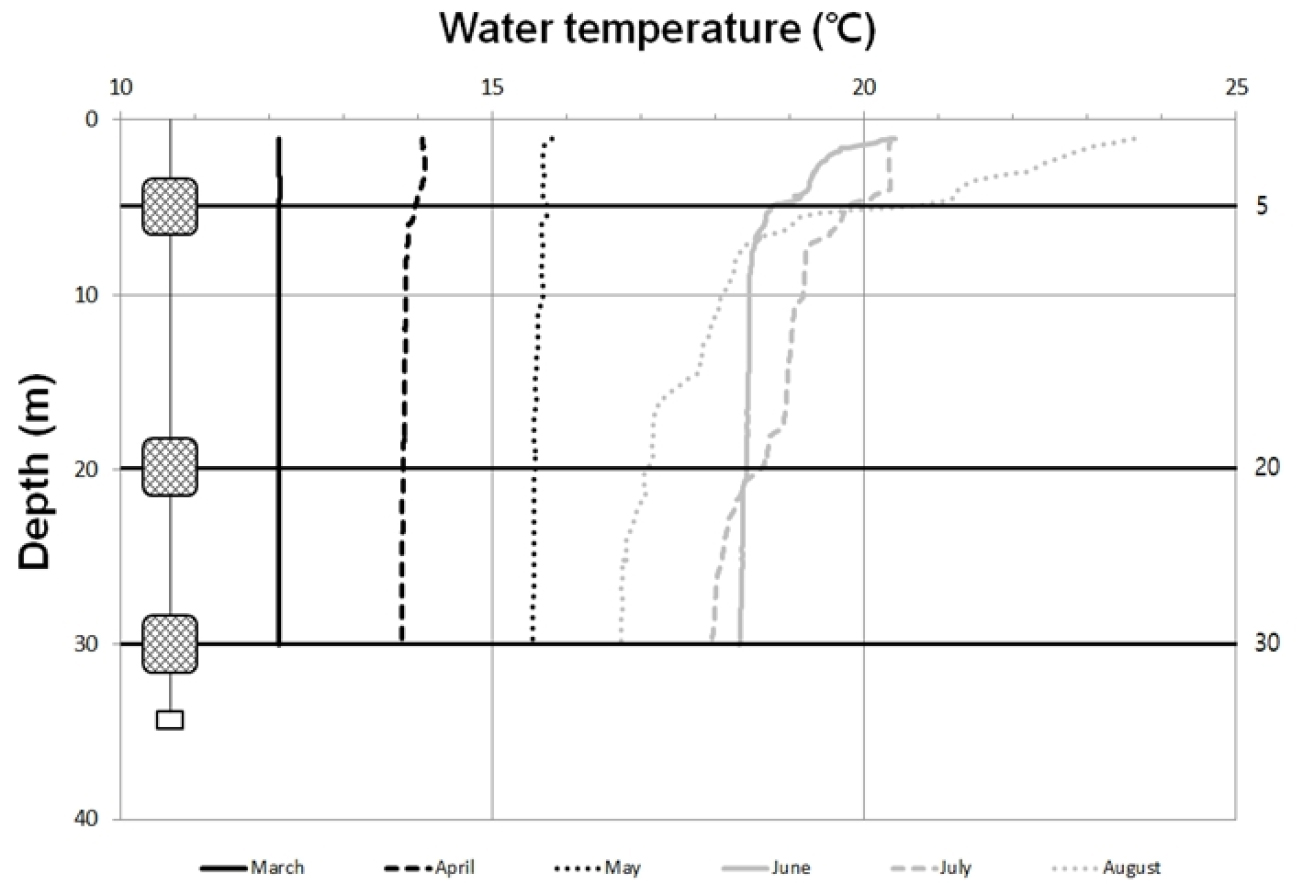
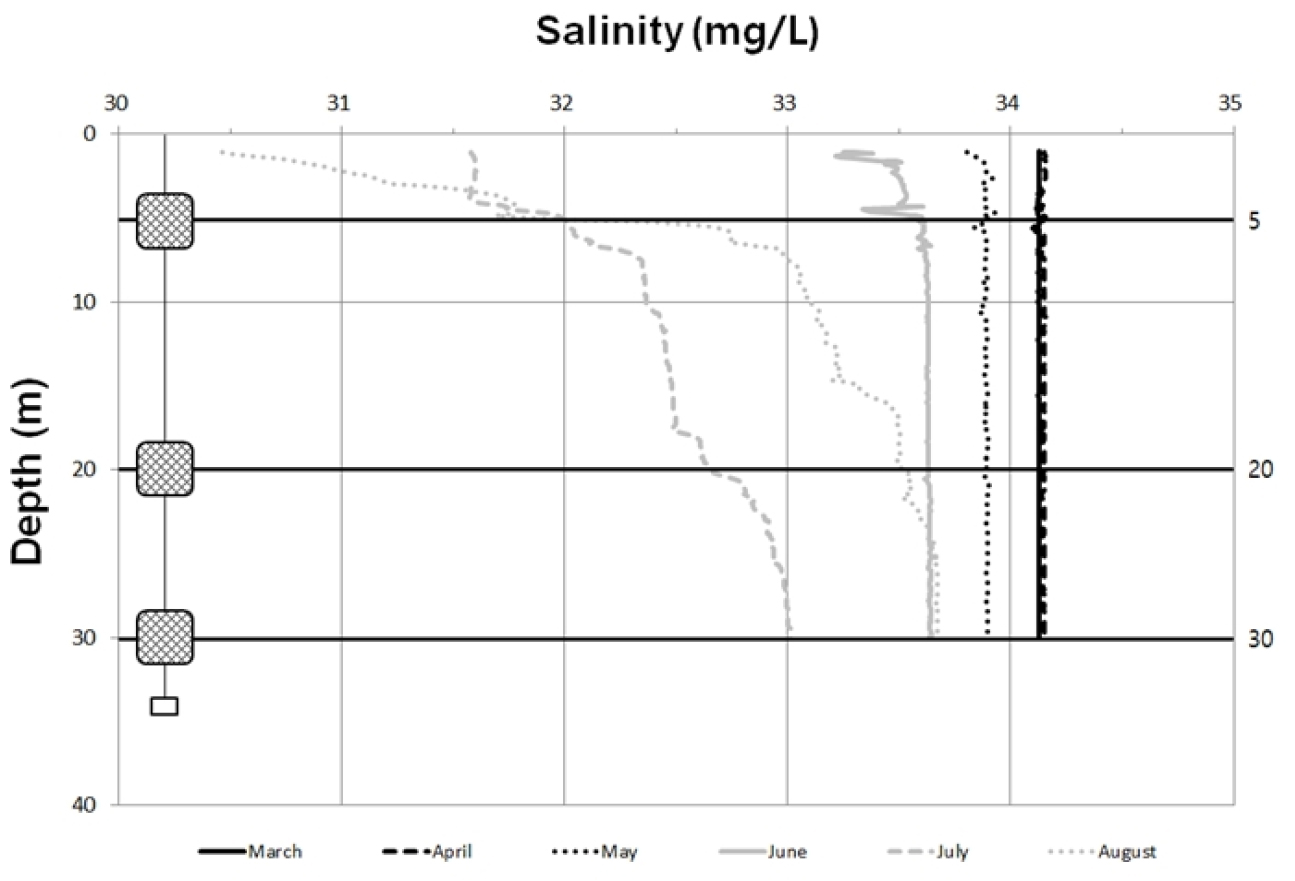
현장에서 계측한 수온 분포는 표층에서 3월(12.2°C)~8월(23.8°C)사이에 10°C 이상 차이를 나타냈고, 30 m 수심에서는 6°C 정도의 수온 차이를 나타냈다(Fig. 2). 수심에 따른 차이는 3월부터 5월 사이에 월별로 유사한 수온을 나타냈으며 약 2°C 전후로 증가하였다. 6월부터는 표층 수온이 20°C 이상 증가하였고, 특히, 8월에는 표층에서 24°C를 나타냈다. 이러한 수온 증가는 주로 수심 5 m와 표층 사이에서 나타났다. 8월에는 오히려 저층 수온이 6월과 7월보다도 1°C 이상 낮았다. 염분은 표층에서 30.5 mg/L(8월)~34.1 mg/L(3월) 범위를 나타냈다(Fig. 3). 3월부터 5월까지 34 mg/L 전후로 월별 또는 수심에 따라 유사하였다. 6월부터는 표층에서 염분도가 점차적으로 감소하였고, 7월과 8월에는 32 mg/L이하를 나타내면서 수직구배에 따라 1 mg/L 이상 차이를 나타냈다. 특히 8월에는 표층에서 30.5 mg/L을 나타냈으며, 30 m 수심에서는 7월보다 높고, 오히려 6월과 유사한 염분도를 나타냈다.
종 조성
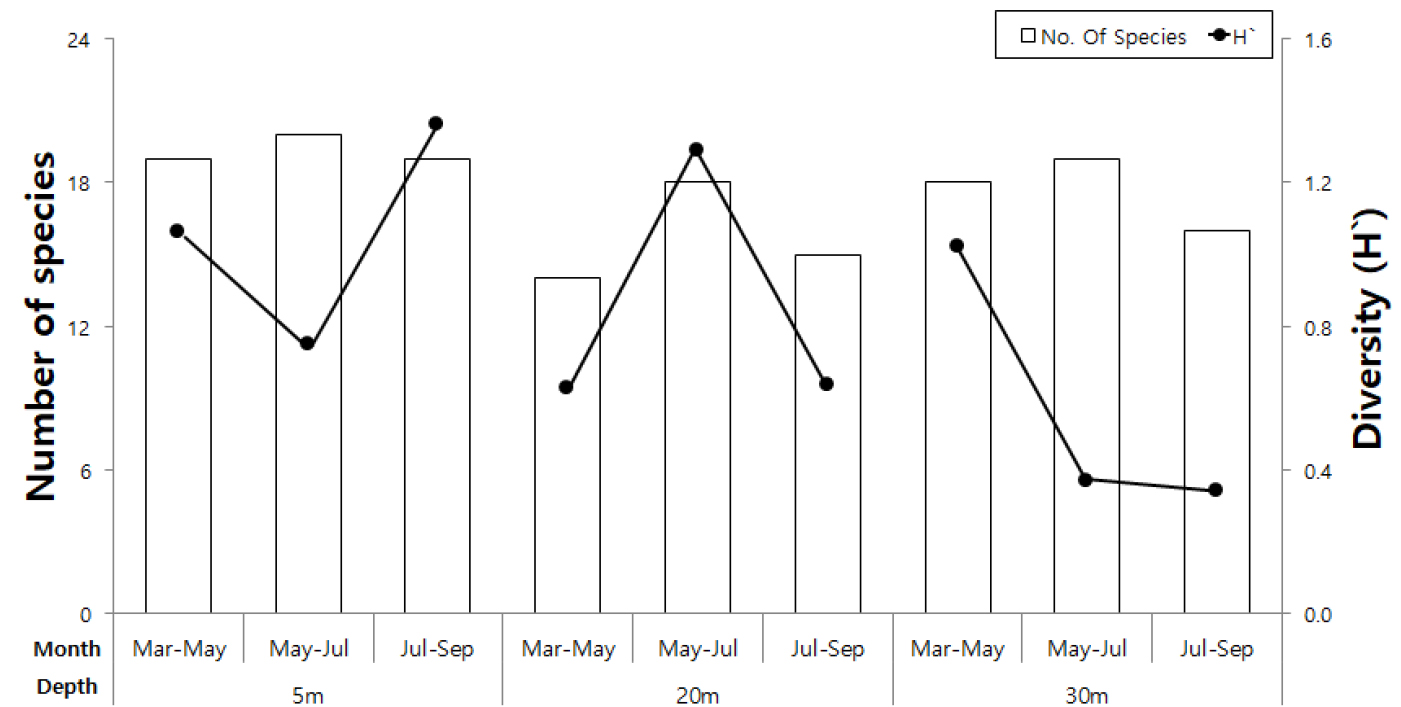
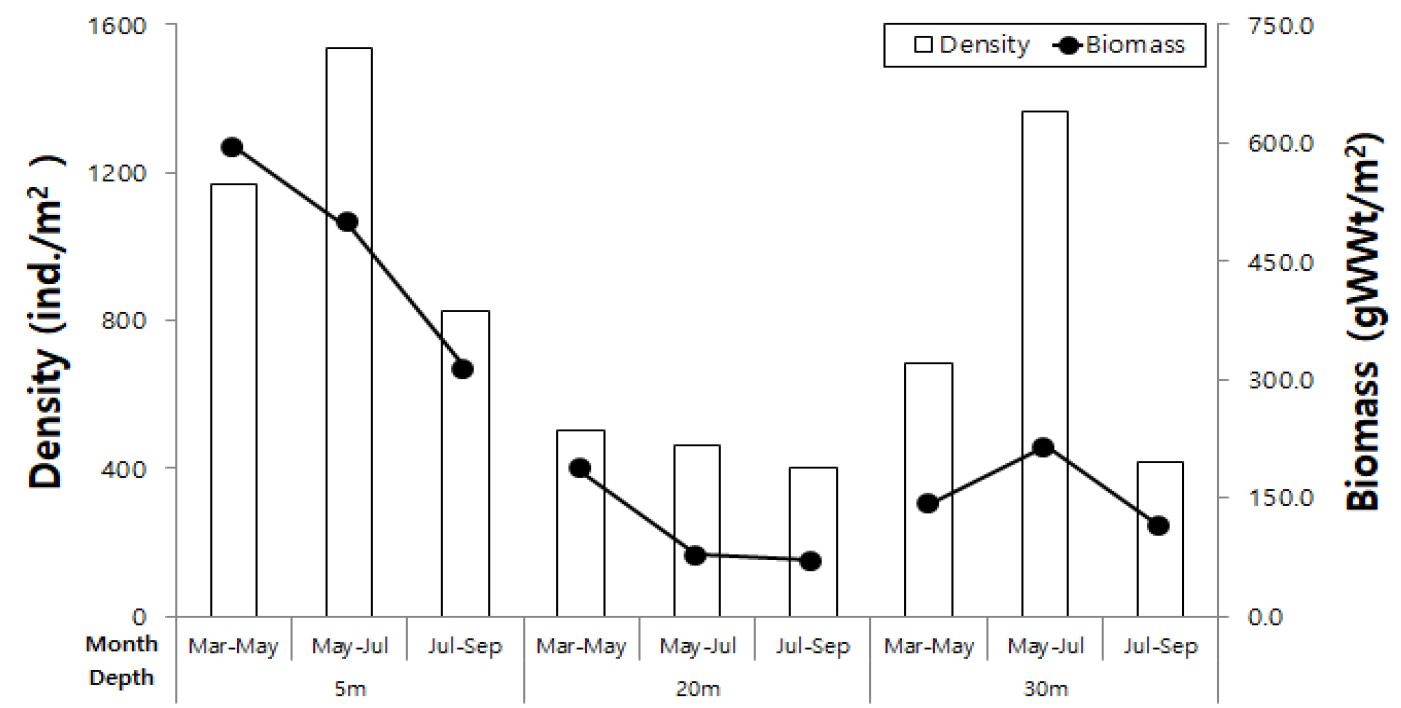
저서동물은 9문, 37종이 출현하였고, 7,361개체, 2126.4 gWWt/m2의 서식밀도와 생체량을 나타냈다(Table 1). 절지동물(Arthropoda)이 14종(37.8%)으로 가장 많은 종 수를 나타냈고, 환형동물(Annelida) 6종(16.2%), 연체동물(Mollusca) 4종(10.8%)이 각각 출현하였다. 서식밀도는 태형동물(Bryozoa)이 2,075개체(28.3%)로 가장 높았으며, 생체량은 곡형동물이 977.6 gWWt/m2(46.0%)로 가장 높았다. 종 수는 수심 5 m에서 30종이 출현하였고 20 m에서는 22종, 30 m에서는 27종이 출현하였다. 수심에 따라 시기별 종 수 변화는 5월~7월에 가장 높았고, 20 m 수심에서 상대적으로 낮게 나타났으며, 시기별 차이도 상대적으로 크게 나타났다(Fig. 4). 다양성 지수는 모든 수심에서 시기에 따라 높은 차이를 나타냈으며 30 m 수심에서 5월 이후 급격하게 낮은 다양도를 나타냈다(Fig. 4). 서식밀도는 5 m 수심에서 상대적으로 높게 나타냈으며, 20 m 수심에서는 가장 낮았다(Fig. 5). 수심 5 m에서 5월~7월 기간에 가장 높은 서식밀도를 나타냈고, 이 시기에는 30 m 수심에서도 높은 서식밀도는 나타냈다. 반면에 계절에 따른 밀도 변화는 20 m 수심에서 가장 적었다. 생체량은 5 m 수심에서 1340.6 gWWt/m2로 가장 높았고, 특히 5월~7월 조사에서 가장 높은 생체량을 나타냈다. 반면에 20 m 수심에서 가장 낮았고, 시기에 따른 편차도 적었다. 30 m 수심에서는 5월~7월에 일시적으로 증가하여, 계절에 따른 생체량의 차이가 크게 나타났다. 5 m, 20 m 수심에서는 시간이 지남에 따라 감소하였다.
Table 1.
Species compositions based on Taxonomic levels
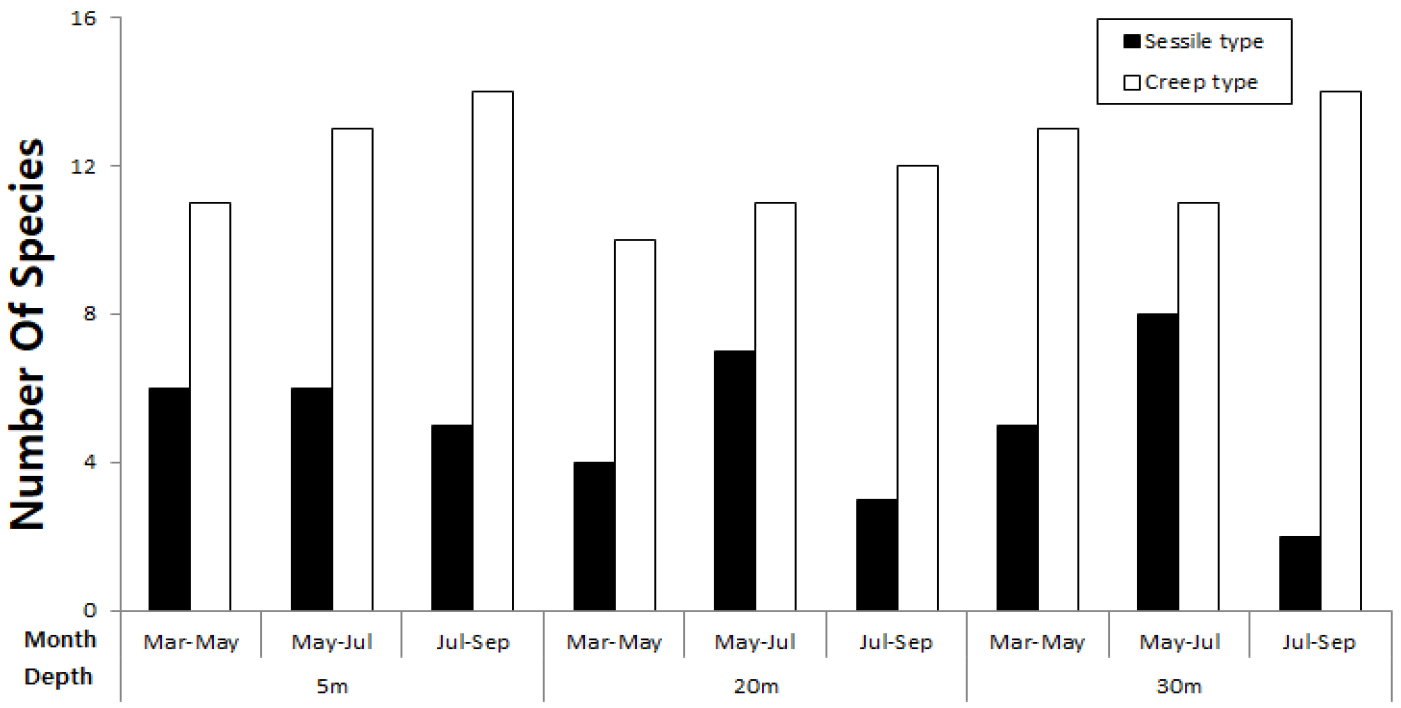
저서동물을 서식형태에 따라 고착성 동물과 포복성 동물로 나누었을 때, 각각 14종, 21종이 포함되었다. 포복성 동물이 고착성 동물보다 모든 수심과 시기에서 높은 서식밀도를 나타냈다(Fig. 6). 고착성 동물은 5월~7월 기간에 종 수가 증가하면서 다른 시기에 비해 높은 종수를 나타냈으며, 특히 30 m 수심에서 가장 높은 종 수를 나타냈다. 7월~9월에는 수심이 깊어질수록 종 수가 감소하였다. 반면에 포복성 동물은 7월~9월에 증가하였고, 수직분포에서는 3월~5월, 7월~9월에는 수심 5 m에서 종수가 많지만 5월~7월에는 수심 30 m에서 상대적으로 높은 종수를 나타냈다.
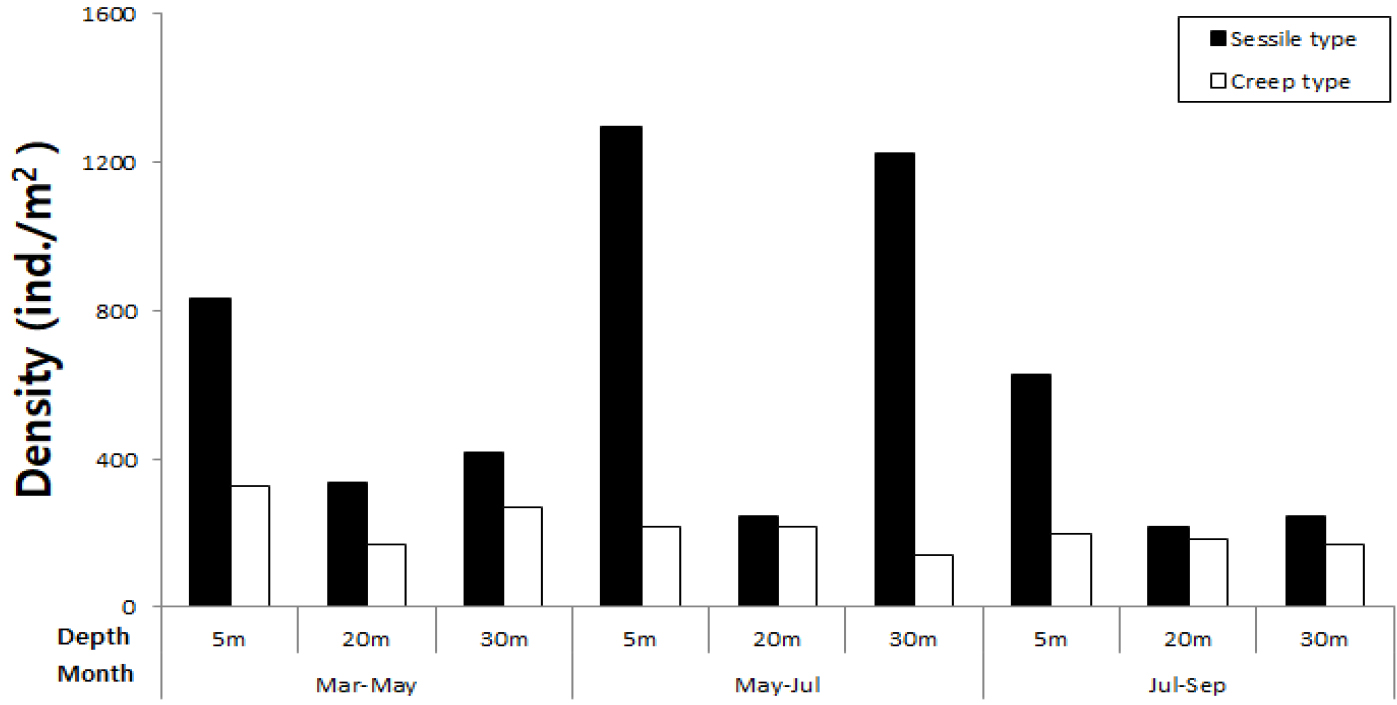
서식밀도에서는 고착성 동물이 포복성 동물에 비해 높게 나타났다(Fig. 7). 모든 시기에서 5 m 수심에서 높은 서식밀도를 나타냈으며, 5월~7월에는 5 m, 30 m 수심에서 대량 가입이 나타났다. 포복성 동물은 모든 시기에 5 m 수심에서 다소 높은 양상을 보였다.
우점종
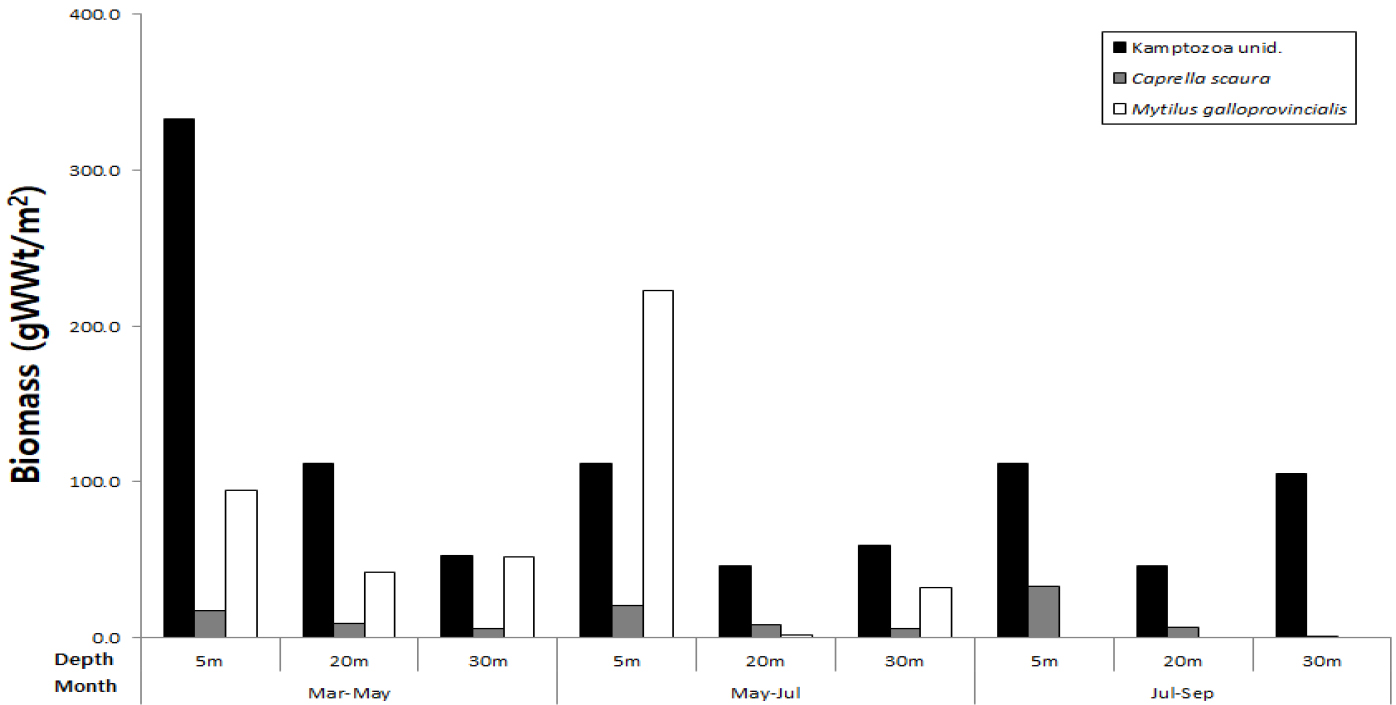
서식밀도와 출현빈도를 고려한 우점종 순위에서 절지동물이 5종 포함되어 가장 우점하는 동물군으로 나타났으며, 곡형동물(Kamptozoa unid.)이 전 기간 동안 모든 정점에서 출현하여 출현빈도와 서식밀도에서 가장 우점한 종 이었다(Table 2). 시기별 분포에서 3~5월 기간에 가장 높은 서식밀도를 나타냈으며, 7월~9월을 제외하고는 5 m 수심에서 상대적으로 높은 밀도를 나타내면서 수심이 깊어질수록 감소하였다(Fig. 8). 바다가시대벌레(Caprella Scaura)의 경우도 시기별로 5 m 수심에서 높은 서식밀도를 나타냈다. 지중해담치(Mytilus galloprovincialis)는 3월~5월, 5월~7월 조사에서는 전 수심에 걸쳐 출현되었으나 7월~9월에는 가입이 나타나지 않았다. 생체량을 기준으로 우점종을 비교한 경우에도 곡형동물이 가장 우세하였으며, 우점종 순위에서는 서식밀도를 기준으로 산정된 우점종과 순위에서 다소 차이를 보였지만 유사한 종들로 나타났다(Table 3). 곡형동물은 생체량에서도 수심 5 m에서 상대적으로 높게 나타났는데, 특히 3월~5월에 수심 5 m에서 가장 높은 생체량을 나타냈다. 지중해담치는 5 m 수심에서 5월~7월이 3월~5월보다 월등히 높은 생체량을 나타냈다(Fig. 9).
Table 2.
Rank of dominant species based on density by Le Bris index
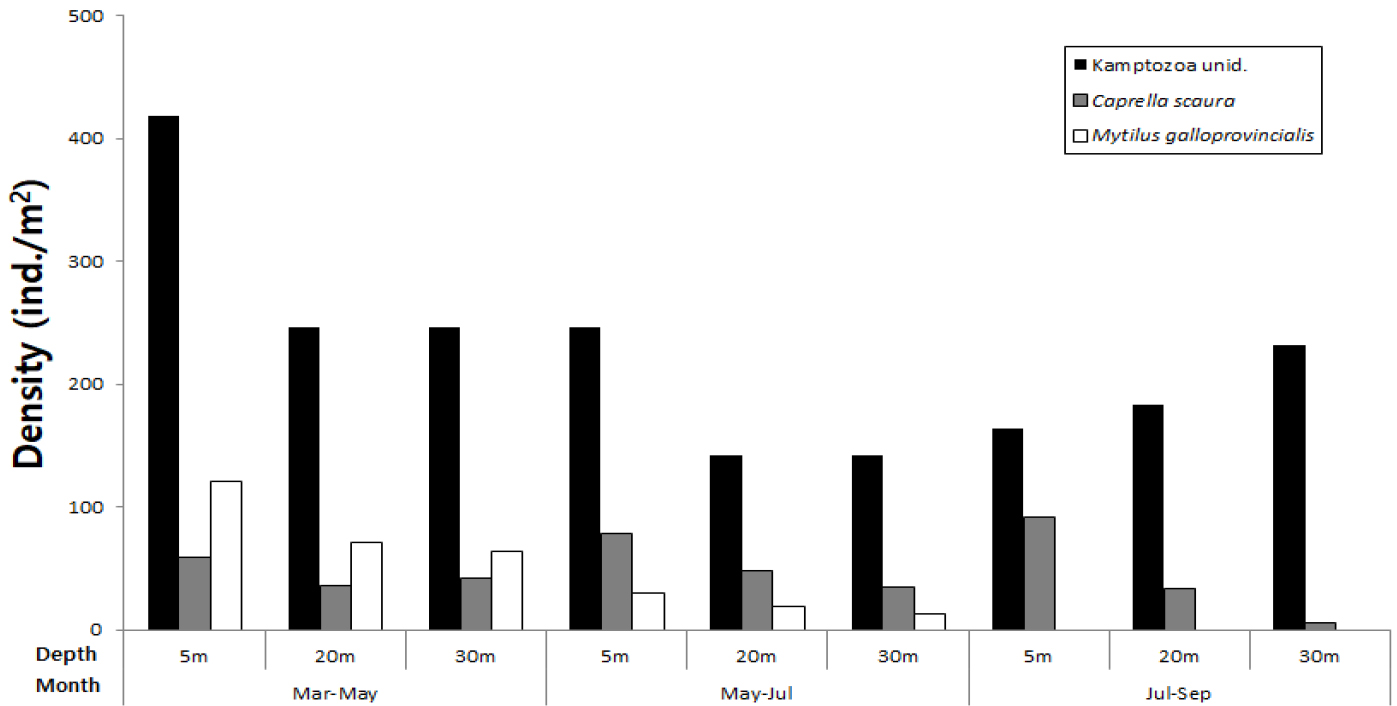
Table 3.
Rank of dominant species based on biomass by Le Bris index
군집 구조
부착판에 가입한 저서동물의 시·공간적 분포는 세 개의 군집으로 구별되었다(Fig. 10). 3월~5월에 5 m와 30 m 수심에서 유사한 종 조성으로 구성된 군집(Group A)으로 구분되었고, 5월~7월과 7월~9월에 5 m 수심이 유사한 군집(Group C)을 형성하였다. 그리고 주로 20 m, 30 m 수심에서 5개 부착판이 유사한 종 조성(Group B)을 형성하였다. 각 군집마다 출현한 종수는 Group A 22종, Group B와 C는 27종이고, 다양도에서는 Group A에서 다소 낮게 나타났으나 군집별로 차이를 보이지 않았다(Table 4). 서식밀도와 생체량에서는 5개 정점이 포함된 Group B에서 오히려 낮았다. 각 그룹을 특정 짓는 기여도가 가장 높은 종으로 Group A는 Bugula neritina, Group B는 Kamptozoa unid., Group C는 Sertularella sp.였다. Kamptozoa unid.와 C. scaura는 Group A, B, C에서 모두 출현하였다.
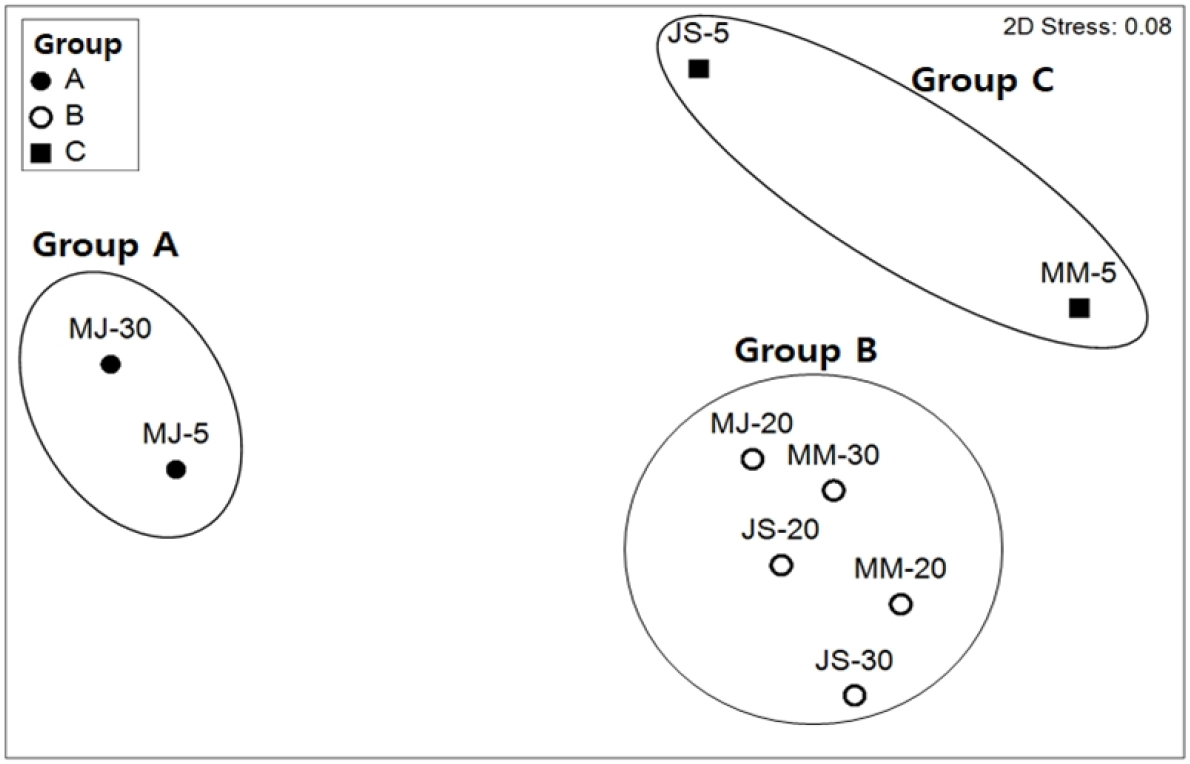
Table 4.
Community characteristics calculated by MDS
각 그룹의 유사도에 영향을 주는 종과 각 그룹을 구분 짓는 종을 파악하기 위한 SIMPER 분석에서 Group A는 태형동물인 B. neritina (71.45%), Group B는 곡형동물인 Kamptozoa unid. (53.30%)와 절지동물인 Jassa slatteryi (10.27%). Group C는 자포동물인 Sertularella sp. (43.00%)가 다른 그룹과 구분되는 영향을 미치는 종으로 나타났다(Table 5).. 각 그룹 간 영향을 미치는 종으로는 Group A와 B는 B. neritina의 영향이 가장 컸고, M. montferradii와 W. subtorquata 등의 영향을 보였다. 군집의 유사도가 낮은 Group A&C에서도 B. neritina의 영향을 나타냈다.
Table 5.
List of contribution species distinguished by SIMPER test
4. 고 찰
여름철 수온 및 염분도 변화
표층의 수온과 염분은 국지적인 태양 복사 및 강우에 따른 직접 영향과 해안으로 유출되는 담수에 의해 간접적 영향을 받는다(이 등 2007; Kang and Jin 1984). 이번 조사에서도 수온의 수직적인 차이가 거의 나타나지 않았고, 6월 이후부터 표층에서 10 m 이내 수심에서 수온이 상승하는 양상을 나타냈다. 여름철이 되면서 상승하는 기온은 표층의 태양 복사열 흡수를 증가시켜 8월에 가장 높은 수온과 수심에 따른 급격한 변화를 나타내지만 8월에는 수심 20 m 이하에서 6월과 7월보다도 낮은 수온이 관측되기도 한다. 하계 제주도 해협과 거문도 인근해역에 걸쳐 형성되는 저층의 냉수괴와 황해저층 냉수가 남해안까지 확장하면서 저층 냉수대의 유입으로 중층 이하의 수온에 일부 영향을 미친다(오 등 2007). 이 등 (2018)에 의하면 여름철 남해 연안수가 고온의 쓰시마 난류 수계와의 사이에서 약 25°C의 수온전선을 형성하지만, 저층을 따라 흐르는 리만해류(북한한류)의 확장과 남풍에 의한 저층 냉수의 용승에 의해, 통영의 경우 여름철에 저층 수온이 표층에 비하여 4~7°C까지 낮아지는 현상이 나타난다고 밝혔다. 현지 어업 인들은 이러한 현상을 ‘청수 유입’이라고 부르기도 한다. 염분도에서도 수심에 의한 차이를 거의 나타내고 있지 않다가 7월과 8월에 표층에서 수직적인 분포에 변화가 최대 3 mg/L 이상 나타났다. 통영은 3월부터 5월 사이에 평균 강수량이 88.8 mm인 반면에 6월부터 8월 사이에는 273.3 mm로 여름철 장마와 잦은 폭우로 인한 담수유입이 조사시기에 측정된 것으로 판단하고 있다(기상청 2019). 여름철 강한 일조와 장마로 인한 높은 수온과 낮은 염분은 부착생물의 가입에 부정적인 영향을 줄 수 있는데 Thiyagarajan et al. (2003)에 의하면 높은 수온(28°C)과 낮은 염분(22 mg/L)은 따개비 Balanus trigonus 유생의 부착률을 33% 미만으로 떨어뜨린다고 보고 하였으며, 전 등(2012)은 굴 Carssostrea gigas 유생이 높은 수온(29°C)와 낮은 염분(20 mg/L)에서 낮은 정착률을 나타냈다고 하였다. 또한 Verween et al. (2007)은 수온이 30°C 이상이거나 염분이 15 mg/L 미만일 때 이매패류 유생 사망률이 가장 높았다고 보고했다. 이번 조사에서 계절에 따른 수온 변화가 군집구조에서는 영향을 미치는 것으로 나타났다. 특히 수심 5 m에서 시기에 따른 상이한 군집이 형성되었다.
종 조성
부착판에 가입한 저서동물 조성에서 수심에 따른 종 조성의 뚜렷한 차이는 나타나지 않았지만 구성하는 분류군이 차이를 보였다. 종 수에서는 포복성 저서동물이 포함된 절지동물이 우세하였고, 서식밀도와 생체량에서는 부착성 저서동물인 태형동물과 곡형동물이 우세하였다. 단위 면적당 서식밀도는 태형동물이 높았고 생체량의 경우는 곡형동물이 높게 나타났다. 태형동물과 곡형동물은 기질에 부착하면서 군체를 이루는 생태를 보이는데, 태형동물은 기질을 수평적으로 점거하면서 개체를 증대시키는 종이고 곡형동물을 동일 부착기에 출아 방식으로 몸체가 수직으로 성장하는 종으로, 국내에서 부착생물의 천이나 종 조성을 조사하는 경우 태형동물이나 곡형동물은 서식밀도를 환산하는 경우가 없고, 개체 수를 기저부 기준으로 파악하거나 기타 동물군으로 취급하고 있다(고 등 2008; 윤 등 2009). 이번 조사가 부착생물에서 서식밀도와 생체량이 우세한 종에 대한 정량적 언급을 시도한 사례가 된다. 모든 수심에서 5월~7월에 높은 종수가 나타났는데 수온이 증가하면서 생물들의 대사 및 생식활동이 활발한 시기이므로 생물들의 종수가 많아진 것으로 판단된다. 남해 장목만에서도 5월 이후 가입되는 생물이 증가하였다(최 등 2011). 평균 다양도 지수는 수심이 깊어질수록 감소하였는데 Michael and Stephen (2007)은 수심이 깊어질수록 생물이 살아가는데 필요한 빛, 온도, 용존산소량이 줄어들고 생존하기에 상대적으로 어려워지기 때문으로 분석하였다. 서식밀도는 수심에 의한 변화보다는 시기에 따른 변화가 뚜렷하게 나타났다. 3월~5월 보다는 수온이 높아지는 5월~7월 사이에 가장 높았으며, 7월~9월 사이에는 오히려 감소하였다. 수온의 증가, 염분도 변화가 크게 변동하지 않는 시기가 부착생물의 성장에 영향을 미치는 것으로 해석할 수 있다. 이러한 양상은 생체량에서도 유사하게 나타났다. 수심에 따른 종 조성의 차이는 주로 5 m 수심에서 상대적으로 높게 나타났는데, 이러한 원인으로는 표층 주변 공간에서 유생의 활동이 가장 활발하여 부착하는 저서동물의 다양성도 높고, 우점하는 생물의 가입 양상도 높게 나타나기 때문으로 추측한다. 인공 부착판에 가입되는 생물들을 한번 정착하면 움직일 수 없는 부착성 생물과 부착판을 자유롭게 이동하는 포복성 생물들로 나누었다. 가입된 생물을 두 분류로 나눈 이유는 포복성 생물은 부착생물군집을 먹이확보, 산란장소, 피난처로 사용하고 부착성 생물은 부착생물군집을 부착을 위한 기질로서 이용하기 때문이다(Michelle et al. 2006).
이번 연구에서도 수심과 조사시기에 따라 포복성 생물의 종수가 부착성 생물보다 우세하였는데, 대부분이 단각류로 바다대벌레류와 옆새우로 구성되었다. Kirsten et al. (2010)은 소형갑각류인 단각류가 부착판 가입이 용의하고, 다른 분류군보다 우세하다고 보고하였다. 또한 수심에 따른 인공기질에서 단각류 서식밀도는 수심이 깊어질수록 감소한다(Gustavo et al. 2013; Michael and Stephen 2007). 서식밀도에서는 포복성 생물보다 부착성 생물이 모든 수심 및 계절에서 우세하였는데 곡형동물류, 히드라충류, 지중해담치의 가입에 의한 영향이다. M. montferrandii는 3월~5월 5 m에서만 출현하였는데 3월~5월에 주로 가입되며 표층에서 서식하는 것으로 예상된다. A. japonica는 3월~7월에 다른 생물들과의 공간경쟁에서 밀려 서식밀도가 낮았지만 7월~9월은 수심 5 m에서 가장 높은 서식밀도가 조사된 것으로 보아 수심이 따뜻해지는 여름철 표층 가까이에서 가장 가입이 활발히 이루어지는 것으로 예상된다.
우점종
우점종은 서식밀도, 생체량, 서식빈도 등 조건에 따라 다양하게 선정하고 있는데, 해당 생태계에서 지속적이기보다는 시기적으로 발생하는 종의 경우 통계적 보정을 통하거나 또는 서식빈도의 비중을 높여서 우점종을 파악하기도 한다. 이번 조사에서도 2개월마다 부착기질을 새롭게 설치한 상황에서 우점종 분포를 파악하기 때문에 서식밀도와 더불어 서식빈도의 비중을 고려한 Le Brix method를 적용하였다. 우점종에서도 시·공간적이 양상을 보이고 있지만, 주요 우점종은 연중 가입이 진행되는 것으로 나타났다. 서식밀도를 비교한 우점종은 곡형동물이 가장 우세하였으며, 전반적으로 포복생물이 높은 비중을 차지하였다. 조사 기간 중에 모든 정점에서 출현한 곡형동물은 해양생물 중에 다양한 종 분화가 진행되지 않아서 종수가 많이 알려져 있지 않은 분류군으로 연안 조류의 흐름이 원활한 곳에 군락을 형성하고 있어서 오손생물로 잘 알려진 동물군이다(Chapman and Carlton 1994). 국내에서는 양식시설을 중심으로 부착기질에 다량 가입하지만 아직 종 분류 및 생태 정보가 거의 밝혀지지 않았다. 미국 동부에서는 연안 암반생태계 교란이 관찰되어 생태교란생물로 지정된 생물군이다(Wasson et al. 2000). 곡형동물은 5 m 수심에서 우세한 서식밀도를 보이고, 여름철 서식양상이 감소하고 3월~5월 시기에 가장 높은 가입량을 보이는 조건을 해석하면 겨울철 주로 가입되는 것으로 예상된다. 우점종에 포복성 저서동물인 단각류가 다수 포함되었다. 이들은 시기적으로 부착생물 군락에 가입하며 머물면서 부착생물 사이에 침전된 유기물을 섭식하거나, 부착기질에 몸을 지탱하고 부유하는 유기물을 포획하기도 한다. 부착 실험구의 경우 2개월인 초기 부착 상황을 나타내어 복잡한 부착기질 생태계 구조가 아닌 부착기질과 초기 부착한 생물이 혼재한 상황인 포복성 생물이 가입하기 용이한 외형 조건을 가지고 있어서 상대적으로 가입량이 높게 나타났다. 특히, 국내에서 진행된 부착생물 연구에서는 포복성 저서동물의 가입에 대한 자료를 배제하고 부착공간에 대한 군집 조성만을 분석하여 부착기질에서 진행되는 종 정보를 알지 못하였다. 이번 조사에서 포복성 저서생물이 시기적으로 연속적인 가입과 높은 생물다양성 등 부착기질에서 종조성의 역할을 언급하였다.
지중해담치는 대서양 연안, 지중해, 홍콩, 일본 남부 등지에 분포하지만 우리나라에 유입되어서 서식하며(성 등 2005), 양식 장비나 해양 구조물에 부착하면서 여름동안 심각한 해안선 기저부에서 오손생물의 역할을 한다. 단년생으로 성장이 매우 빨라서 다른 부착성 생물과 공간경쟁에서 우위를 차지하지만, 급격한 성장으로 종간 공간 경쟁과 여름철 파도와 태풍의 영향으로 집단 이탈하며, 군집의 틈새 공간이나 패각 표면에는 다양한 동물의 서식공간을 제공한다(Chintiroglou et al. 2004). 포복성 저서동물은 지중해담치 군집의 복잡한 기질에서 먹이나 찌꺼기들을 찾아다니며(Çinar et al. 2007), 지중해담치의 패각은 다른 부착생물에게 서식공간을 제공한다(Thiel and Ullrich 2002). 이번 조사에서도 3월에 모든 수심에서 우세한 가입양상을 보이다가 5월에는 감소하고, 7월에는 전혀 가입되지 않았다. 또한 유생가입이 시간이 지날수록 저층부터 감소하는 양상을 파악하였다.
군집특성
수심과 시기별 부착생물 가입양상에 대한 유사도를 비교한 결과 수심이 군집조성에 영향을 미치는 것으로 나타났다. 부착생물의 분포와 군집구조 형성에서 수심은 중요한 영향을 미치는 인자이며(Witman 1985; Underwood et al. 1991; Gallahar and Kingsford 1992; Witman and Dayton 2001), 서식공간(Underwood and Denley 1984; Underwood and Petraitis 1993)과 시기(Hutchinson and Williams 2001) 등도 중요하게 작용한다. 군집구조는 5월~7월 수심 30 m 조건을 제외하고는 수심 20, 30 m에서 종조성이 유사한 것으로 나타났다, 다만 5월~7월의 수심 5 m와 30 m에서 유사한 군집을 나타낸 것은 당시 공간에서 우점종은 곡형동물이었지만 태형동물인 B. neritina의 가입이 영향을 미친 것으로 나타났다. 동일시기에 수심 20 m에서도 소수의 B. neritina이 가입하였지만 상대적으로 적은 서식밀도가 출현하였다. 이러한 군집구조의 양상을 고려하면, 대상 해역에서 부착생물의 시기에 따른 가입양상은 수심에 따라 차이를 나타내며, 이러한 결과는 표층을 중심으로 진행된 기존 연구에서는 인식할 수 없는 결과이다. 현재까지 수심 20 m 이후 공간을 활용하는 방안은 매우 적지만, 수하식 양식이나 침설식 양식등에 중요한 정보를 제공할 수 있다. 수심 5 m는 시기에 따라 우점종의 가입량 변동이 발생하고, 포복성 저서동물의 가입이 다른 수심에 비해 원활하였다. 수심 20 m와 30 m의 경우 시기적으로 가입되는 종이 상이하지만 평균 서식밀도는 유사하다. 군집구조에 중요하게 작용하는 종은 곡형동물, 태형동물, 히드라충류 등 주로 부착성 저서동물이며, 이러한 결과는 각 군집이 종조성에 의한 차이보다는 특정 종이 공간적 선택성에 의해 대량 가입유무로 인해 군집구조가 형성된다고 해석할 수 있다. SIMPER 분석에서도, 태형동물인 B. neritina과 곡형동물인 Kamptozoa unid.이 모든 시기에서 수심 20 m와 30 m에서 출현하고, 수온 변동이 관측된 5 m에 비해 안정된 서식양상을 나타냈다.
