

1. 서 론
2. 재료 및 방법
연구지역 및 시료 채취
시료 분석
통계 분석
3. 결과 및 고찰
대체 질소원을 필요로 하는 질소 제한 시스템
보편적이고 유용한 질소원으로서의 요소
생태계 상태의 전이(State Transition)와 적조 발생의 동인
적조 발생에 따른 자생적 유기물 생성과 생지화학적 되먹임
4. 결 론
1. 서 론
연안 생태계는 육상과 해양이 만나는 경계 영역으로, 지구 생지화학적 순환에서 일정한 역할을 수행한다(Walsh 1991). 산업화와 인구 증가로 인해 다량의 영양염이 연안으로 유입되면서 부영양화(eutrophication)가 전 세계적으로 진행되고 있다(Schlünz and Schneider 2000; Chen and Borges 2009). 과도한 질소(N)와 인(P) 공급은 식물플랑크톤의 급격한 증식, 즉 유해성 적조(HABs)를 유발할 수 있으며, 이는 용존산소 고갈, 수산자원 폐사 등의 환경 문제로 이어진다(Anderson et al. 2002; Rabalais 2002).
해양의 신생산(new production)은 주로 질산염(NO3-)과 같은 용존무기질소(DIN)에 의해 지배된다. 그러나 많은 연안, 특히 온대 해역은 여름철에 DIN이 빠르게 고갈되어 질소 제한(N-limitation) 상태가 된다(Conley et al. 2009). 이러한 환경에서 식물플랑크톤은 용존유기질소(DON)를 대체 질소원으로 활용하므로, DON의 양과 질이 일차생산력과 군집 구조를 결정하는 요인이 된다(Bronk 2002).
DON의 구성 성분 중 하나인 요소(Urea, CO(NH2)2)는 식물플랑크톤이 이용 가능한 질소원이다(Glibert et al. 2006; Solomon et al. 2010). 요소는 생물 배설물이나 유기물 분해를 통해 자연적으로 생성될 뿐만 아니라, 농업용 비료 등 인위적 경로를 통해서도 연안으로 공급된다(Glibert et al. 2006). 식물플랑크톤은 urease 효소를 이용해 요소를 암모늄(NH4+)으로 전환하여 질소원으로 사용하며, 이는 질산염을 이용하는 것보다 에너지 효율이 높다(Fan et al. 2018). 일부 연구에서는 특정 조건에서 요소가 전체 질소 흡수의 50% 이상을 차지하며, 일부 유해 와편모조류의 성장을 촉진한다고 보고되었다(Glibert et al. 1991; Berg et al. 1997).
복잡한 해안선과 조밀한 양식장이 분포한 한국 남해안은 여름철 고수온과 장마철 육상 오염원 유입이 겹치면서 매년 유해성 적조가 반복적으로 발생한다(Lee 2006; Lee et al. 2013). 남해안의 적조 발생 기작 연구는 주로 DIN, 수온, 염분 등 전통적인 환경 요인에 집중되었으며, DON, 특히 요소의 역할에 대한 평가는 부족했다.
따라서 본 연구는 여름철 질소 제한이 심화되는 한국 남해안에서 요소의 시공간적 분포와 동태를 파악하고, 식물플랑크톤 대발생과의 연관성을 규명하고자 하였다. 이를 위해 (1) 여름철(6–8월) 표층수의 영양염 분포를 분석하고, (2) DON 내 요소의 비율을 통해 생체이용가능성을 평가하며, (3) 다변량 통계 분석으로 적조 발생과 환경 요인 간의 관계를 분석하였다.
2. 재료 및 방법
연구지역 및 시료 채취
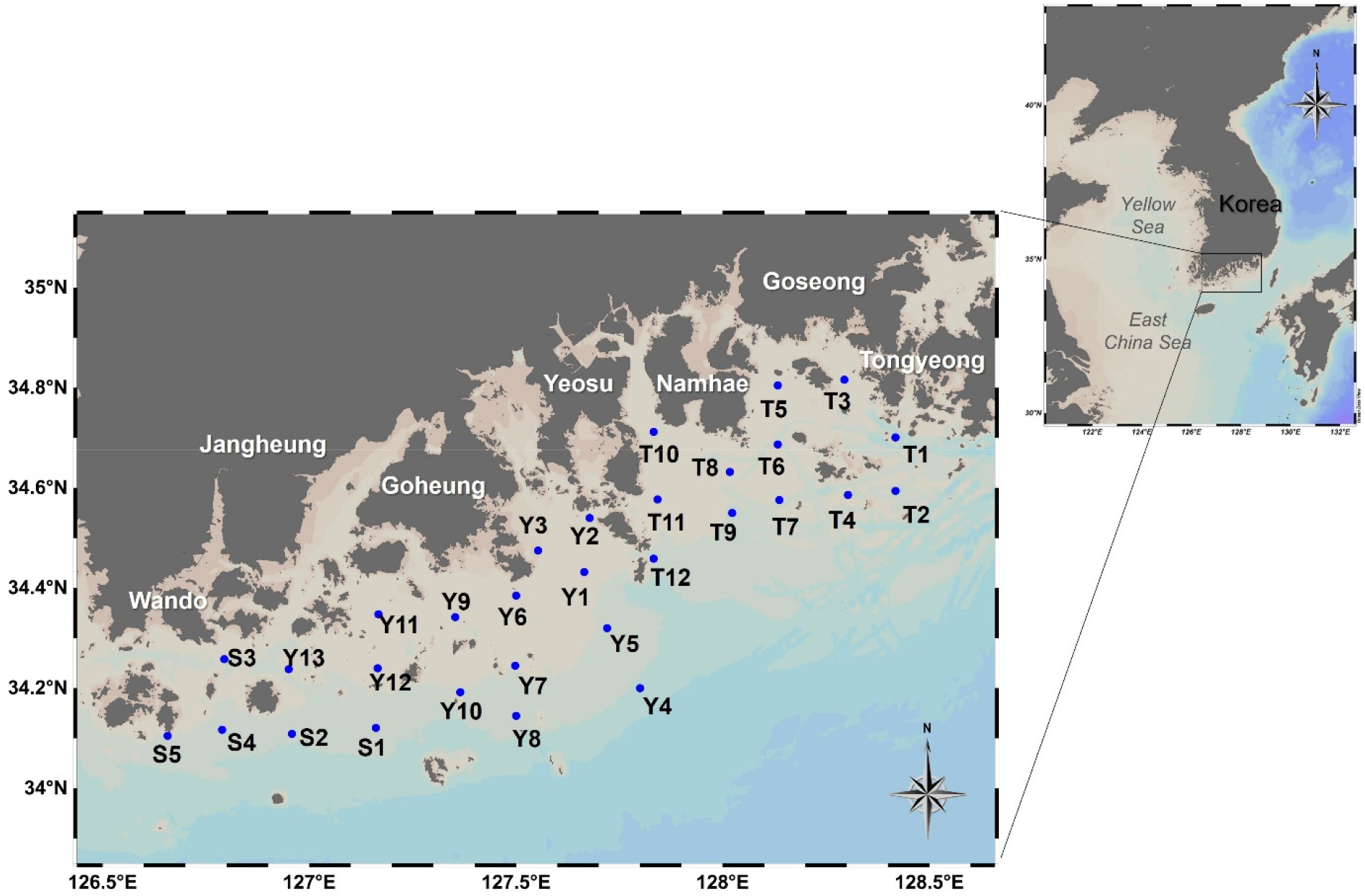
본 연구는 한국 남해안의 완도 동쪽 해역부터 통영 서쪽 해역에 이르는 연안을 대상으로 수행되었다(Fig. 1). 이 해역은 담수의 영향을 받는 내만 정점부터 외양수의 영향을 받는 외해 정점까지 이어지는 뚜렷한 염분 및 영양염 구배를 포함한다. 현장 조사는 여름철 계절적 환경 변화를 포괄하기 위해, 2023년 6월 19–23일(장마 전), 7월 17–21일(장마 중), 8월 13–18일(장마 후/고수온기) 총 3회에 걸쳐 국립수산과학원 수산과학조사선을 이용하여 수행되었다. 각 조사 정점에서 표층 해수(수심 약 0.5 m)를 10% HCl로 산 세척 후 초순수(Milli-Q)로 3회 이상 헹군 폴리카보네이트(PC) 채수병을 이용하여 채취하였다. 채취된 시료는 500°C에서 5시간 이상 회화(pre- combusted)하여 유기물을 완전히 제거한 Glass Fiber Filter (GF/F, Whatman, 공칭 공극 0.7 µm)를 이용하여 선상에서 즉시 감압 여과하였다. 여과액과 필터는 분석 항목에 따라 각각 별도로 처리하여 냉동 또는 냉장 보관하였다.
시료 분석
수온 및 염분은 현장에서 CTD (Conductivity-Temperature-Depth) 센서(SBE 19plus, Sea-Bird Electronics)를 이용하여 수심별 수온과 염분을 연속적으로 측정하였다. 무기 영양염(DIN, DIP, DSi)은 여과액 15 mL를 폴리프로필렌 코니컬 튜브에 분취하여 분석 전까지 -20°C에서 급속 냉동 보관하였다. 실험실에서 해동 후, 5채널 영양염 자동분석기(QuAAtro39, SEAL Analytical)를 이용하여 표준 비색법에 따라 분석하였다(Grasshoff et al. 1983). 질산염(NO3-)과 아질산염(NO2-)은 카드뮴-구리 환원 칼럼을 통과시킨 후 술파닐아마이드(sulfanilamide)와 반응시켜 측정하였고, 인산염(DIP, PO43-)은 몰리브덴산-아스코르브산법으로, 규산염(DSi, Si(OH)4)은 몰리브덴산-청법으로 분석하였다. 암모늄(NH4+)은 페놀-하이포아염소산염법(o-phthalaldehyde method)으로 분석하였다. DIN은 NO3-, NO2-, NH4+ 농도의 합으로 정의하였다. 분석의 정확도와 정밀도를 확보하기 위해 표준물질(RMNS, Kanso)을 사용하였으며, 검출 한계는 NO3- 0.02 µM, NO2- 0.01 µM, NH4+ 0.05 µM, DIP 0.01 µM, DSi 0.1 µM 이하였다. 분석 정밀도는 모든 분석 항목에서 5% 이내였다.
요소 분석을 위한 여과액 50 mL는 폴리프로필렌(PP) conical tube에 담아 -20°C에서 냉동 보관하였다. 분석은 diacetyl monoxime을 이용한 수동 비색법(Goeyens et al. 1998)에 기반하여 수행하였다. 요소의 검출 한계는 0.05 µM이었고, 분석 정밀도는 1% 이내를 유지하였다. 해동된 시료에 시약을 첨가하고 85°C에서 30분간 가열한 후, 실온에서 냉각시켜 생성된 분홍색 복합체의 흡광도를 10 cm 석영 셀을 사용하여 분광광도계(Spectrophotometer)로 520 nm 파장에서 측정하였다. DON 농도는 고온촉매산화법으로 측정한 총용존질소(TDN)에서 상기 분석된 DIN 농도를 감하여 산출하였다. 용존유기탄소(DOC) 와 총용존질소(TDN) 분석용 여과액 약 20 mL를 pre-combusted borosilicate glass ampoule에 담고, 미생물 활동 억제를 위해 고순도 6M HCl 20 µL를 첨가하여 pH를 2 이하로 조절하였다. ampoule 입구를 토치로 녹여 완전히 밀봉한 후, 분석 전까지 암소의 실온에 보관하였다. DOC 및 TDN 농도는 고온촉매산화법(HTCO, 680°C)을 이용하는 총유기탄소/질소 분석기(Shimadzu, TOC-VCPH/ TNM-1)로 측정하였다. 분석의 정확도는 국제 표준물질(Deep Sea Reference, Hansell Lab, University of Miami)을 이용하여 검증하였으며, 분석 오차는 3% 이내였다.
형광용존유기물(FDOM)는 여과 시료를 pre-combusted 암갈색 유리병에 담아 분석 전까지 4°C에서 냉장 보관하였다. 형광광도계(SCINCO, FluoroMate FS-2)를 이용하여 3차원 Excitation-Emission Matrix (EEMs)를 획득하였다. 들뜸 파장(Excitation)은 250–400 nm (5 nm 간격), 방출 파장(Emission)은 250–550 nm (2 nm 간격) 범위에서 측정하였다. 측정된 형광 강도는 Milli-Q 물을 이용한 블랭크 보정, 내부 필터 효과(inner-filter effect) 보정, 라만 산란 보정을 거쳤으며, Quinine Sulfate 등가 단위(QSU)로 표준화하였다. EEMs 데이터셋은 MATLAB 환경에서 PARAFAC (Parallel Factor Analysis) 모델링(DOMFluor 툴박스)을 통해 개별 형광 성분으로 분리하였고, split-half validation과 잔차 분석을 통해 모델의 안정성과 유효성을 검증하였다.
통계 분석
모든 통계 분석은 R-Studio (ver. 4.2.1)와 SPSS (ver. 19.0)를 이용하여 수행하였다. 월별 환경 변수 간의 통계적 유의성 검증을 위해, 데이터의 정규성(Shapiro-Wilk test) 및 등분산성(Levene’s test)을 확인한 후, 일원배치 분산분석(One-way ANOVA)과 Tukey's HSD 사후 검증을 적용하였다. 정규성을 만족하지 않는 데이터의 경우, 비모수 검정인 Kruskal-Wallis test를 실시하였다. 유의수준은 p < 0.05로 설정하였다. 적조 발생 시기의 다차원적 환경 특성을 규명하기 위해 주성분 분석(PCA)을 실시하였다. 분석에 사용된 5개 변수(수온, 염분, 요소, DIN, DIP)는 단위 차이로 인한 왜곡을 방지하기 위해 각 변수의 최대값으로 나누는 정규화(normalization) 과정을 거친 후 분석에 사용하였다(Legendre and Legendre 2012).
3. 결과 및 고찰
대체 질소원을 필요로 하는 질소 제한 시스템
조사 기간 동안 연구 해역의 물리·화학적 환경은 뚜렷한 계절적 변화를 보였다. 표층 수온은 6월(평균 20.8 ± 1.1°C)에서 8월(평균 26.8 ± 0.9°C)로 상승하며 여름철 성층 발달을 위한 조건을 형성하였다. 반면, 염분은 6월(평균 33.8 ± 0.3)에 가장 높았고, 7월에 내만 정점을 중심으로 30.5까지 하락한 후 8월에 다소 회복되어(평균 32.1 ± 0.8), 7월의 강우가 육상 기원 담수의 대량 유입을 야기했음을 보여준다.
DIN 농도는 6월(0.0–4.4 µM) 이후, 집중 강우와 함께 육상 오염원이 유입된 7월에서 0.1–8.3 µM로 최댓값을 기록하였다(Fig. 2b). 특히 외측 정점에서 관찰된 높은 DIN 농도는 육상 유입수의 장거리 확산과 더불어, 담수 유입으로 강화된 성층과 국지적 해수 순환이 영양염 집적에 복합적으로 기여했을 가능성을 시사한다. 이후 8월에서는 0.2–0.7 µM로 대부분의 해역에서 고갈되었는데(Fig. 2c), 이는 고수온 환경에서 식물플랑크톤의 흡수 활동이 일어났음을 시사한다. DIP 농도 역시 유사한 경향을 보였으며, 8월에는 평균 0.03 ± 0.01 µM까지 감소했다(Fig. 2f).
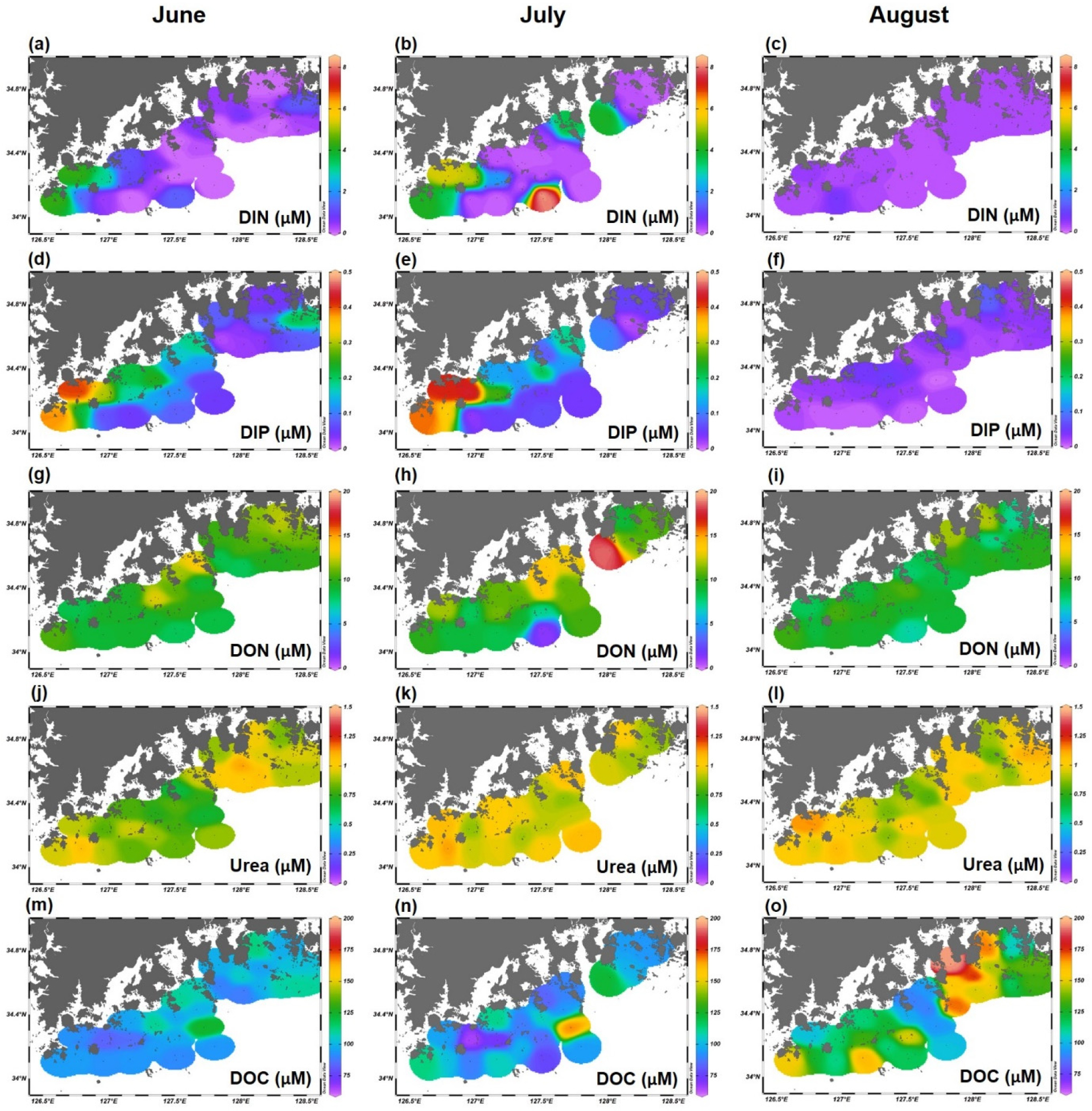
Fig. 2.
Surface water concentrations of Dissolved Inorganic Nitrogen (DIN), Dissolved Inorganic Phosphate (DIP), Dissolved Organic Nitrogen (DON), Urea, and Dissolved Organic Carbon (DOC) in the southern coastal waters of Korea during June (a, d, g, j, m), July (b, e, h, k, n), and August (c, f, i, l, o) 2023
조사 기간의 주요 특징은 연구 해역이 기본적으로 뚜렷한 질소 제한(Nitrogen Limitation) 환경이라는 점이다. 7월의 영양염 펄스로 일부 정점에서 일시적으로 완화되었으나, 전체 조사 정점의 평균 DIN:DIP 비는 11.2 ± 3.5로, Redfield 비(16)보다 유의하게 낮았다(t-test, p < 0.01) (Fig. 3). 이러한 지속적인 질소 결핍 환경은 남해안 생태계의 일반적인 특성으로 알려져 있으며(Conley et al. 2009), 식물플랑크톤이 DIN 이외의 대체 질소원을 필요로 하는 생태학적 압력으로 작용한다.
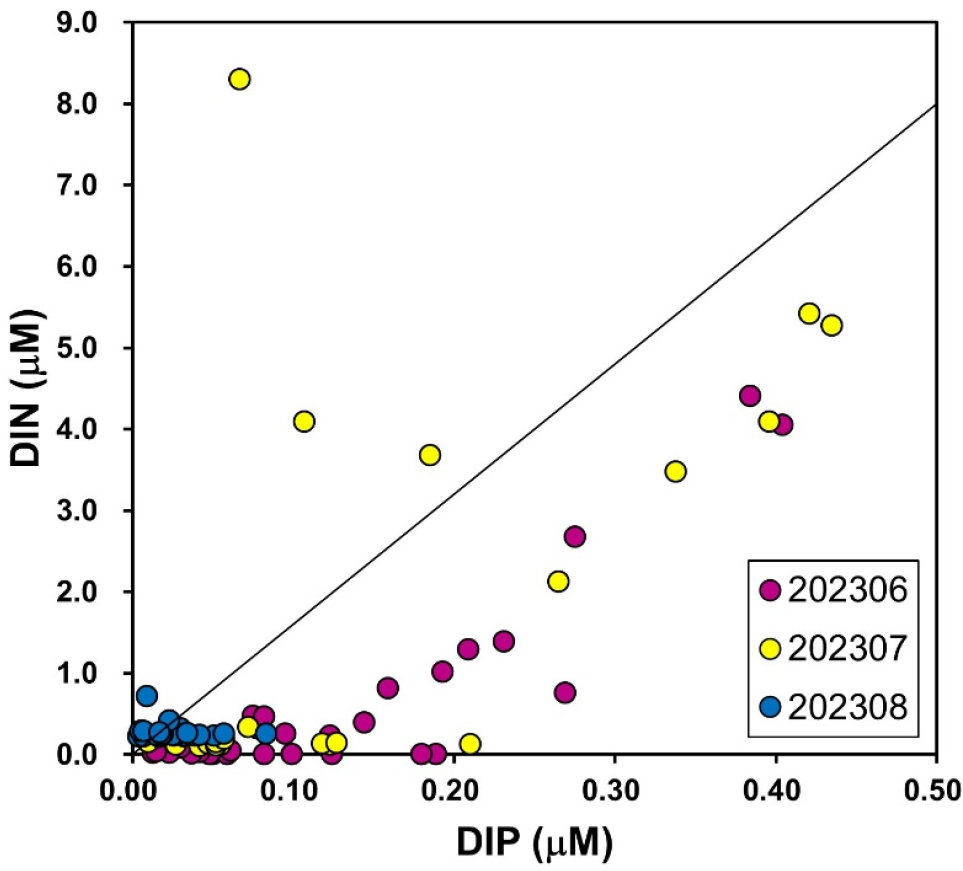
Fig. 3.
Relationship between Dissolved Inorganic Nitrogen (DIN) and Dissolved Inorganic Phosphate (DIP) concentrations at the surface in the southern coastal waters of Korea. Data points are color-coded by month: June (purple), July (yellow), and August (blue). The solid line represents the Redfield ratio (N:P = 16:1)
보편적이고 유용한 질소원으로서의 요소
DON과 요소 농도는 DIN 농도의 최댓값 및 염분의 최솟값을 보인 7월에 가장 높게 나타나, 장마철 육상 유입이 유기물의 주요 공급원임을 시사한다(Fig. 2). DON과 요소 농도는 염분과 음의 상관관계(각각 r = -0.68, r = -0.75, p < 0.001)를 보여 담수 유입이 이들의 주요 공급 경로임을 뒷받침했다. 전체 조사 정점의 평균 요소 농도(0.9 ± 0.2 µM, 범위 0.6–1.6 µM)는 미국 체서피크만(Chesapeake Bay)에서 보고된 여름철 요소 농도(0.2–1.5 µM)와 유사한 수준이었다(Lomas et al. 2002).
본 연구에서 DON 중 요소가 차지하는 비율(Urea: DON)은 평균 0.11 ± 0.09로, 생체이용가능성의 임계치로 알려진 0.05를 초과했다. 이는 DON의 생체이용가능성이 높고, 적조 발생과 관련이 있다는 선행 연구와 일치한다(Zhang et al. 2020). 또한, 요소는 암모늄 농도가 높은 환경에서도 일부 와편모조류에 의해 섭취될 수 있어(Glibert et al. 1991), 특정 적조생물의 경쟁적 우위를 확보하게 하여 적조 발생의 직접적인 원인으로 작용할 수 있는 잠재력을 지닌다.
생태계 상태의 전이(State Transition)와 적조 발생의 동인
주성분 분석(PCA) 결과, 여름철 남해안 생태계가 세 가지의 생태학적 상태(ecological state) 간의 전이(transition)가 나타났다(Fig. 4). 6월의 연구 해역은 고염분-빈영양 유사 상태(Oligotrophic-like state)로 구분된다. 이 시기는 외양수의 영향으로 인해 상대적으로 염분이 높고 영양염 농도가 낮아 생물 활동이 제한된 준평형 상태(quasi-equilibrium state)를 보였다.
Fig. 4.
Principal Component Analysis (PCA) of environmental variables (temperature, salinity, urea, DIN, and DIP) for samples collected in June (blue dots), July (orange dots), and August (green dots) 2023. The percentage of variance explained by each principal component is shown in parentheses on the axes
그러나 여름 몬순과 연계된 집중 강우라는 외부 교란(external perturbation)은 시스템을 7월의 영양염 펄스-주도 상태(Nutrient Pulse-driven state)로 전환시켰다. PCA 로딩 플롯에서 7월 시료군이 ‘요소’와 ‘DIN’ 축 방향으로 양의 상관관계를 보이며 군집하는 것은 이러한 상태 전이가 육상 기원 영양염, 특히 요소의 유입에 의해 촉발되었음을 시사한다. 이 영양염 펄스는 일시적으로 질소 제한을 완화하여, 이 해역에서 역사적으로 반복 발생해 온 Cochlodinium polykrikoides와 같은 유해 와편모조류의 대규모 증식을 유발할 수 있는 요인으로 작용한다(Lee 2006; Lee et al. 2013). 실제로, 2023년 8월에는 과거와 같은 대규모 Cochlodinium 적조가 보고되지 않았으나(NIFS 2023), 본 연구에서 관찰된 높은 요소 농도와 DIN 고갈 환경은 적조 발생의 전제 조건을 형성하기에 충분했다. 대발생 이후, 시스템은 8월의 고수온-자생적 상태(High Temperature-Autotrophic state)로 전이되었다. 이 단계에서는 시스템 제어 동력이 외부의 영양염 유입(allochthonous input)에서 식물플랑크톤 자체의 생산 및 분해와 같은 내부 생물학적 과정(autochthonous processes)으로 전환된다.
이러한 분석은 한국 남해안의 여름철 유해성 조류 대발생이 불연속적인 생태계 상태 전이를 통해 진행됨을 보여준다. 특히 몬순 시기의 집중 강우를 통해 유입되는 요소는 이러한 연쇄 반응을 개시하는 촉발 인자(key triggering factor)로 작용한다.
적조 발생에 따른 자생적 유기물 생성과 생지화학적 되먹임
식물플랑크톤의 대규모 증식은 생태계의 유기물 풀(pool)과 순환 과정에 영향을 미친다. 본 연구에서 8월에 관찰된 DOC 농도의 증가(Fig. 2o)는 적조 발생 이후 나타날 수 있는 뚜렷한 생지화학적 변화 중 하나이다. 이러한 해석을 뒷받침할 수 있는 증거는 DOC 농도와 단백질 계열 FDOM (tryptophan-like, Peak T) 간의 유의미한 양의 상관관계(r2 = 0.52, p < 0.001)에서 찾을 수 있다(Fig. 5). 단백질 계열 유기물은 생물 활동에 의해 생성된 ‘신선한(fresh)’ 자생적(autochthonous) 유기물의 지시자로 알려져 있다(Coble 1996; Stedmon and Markager 2005). 따라서 8월에 관찰된 DOC의 증가와 단백질 계열 FDOM의 증가가 동반되었다는 사실은 이 시기에 축적된 유기물이 주로 식물플랑크톤의 세포 외 유기물 방출(exudation) 및 세포 사멸(senescence and mortality) 과정에서 유래했음을 시사한다. 8월에 관찰된 DOC의 증가는 DON의 감소뿐만으로는 설명되지 않을 수 있는데, 이는 대규모로 증식한 식물플랑크톤이 세포 외 유기물 방출 및 사멸 과정을 통해 질소보다 탄소가 풍부한 유기물을 대량으로 방출했기 때문으로 판단된다.

‘외부 영양염 유입(7월) → 적조 발생 → 자생적 유기물 생성 및 축적(8월)’으로 이어지는 일련의 과정은 외부의 영양염 펄스가 시스템 내부의 탄소 순환을 어떻게 변화시키는지를 보여준다. 즉, 7월에 유입된 요소와 DIN은 질소원으로서 식물플랑크톤의 성장을 직접적으로 촉진했을 뿐만 아니라, 그 결과로써 8월에는 생태계 내 탄소 저장고인 DOC의 양을 크게 증폭시키는 간접적인 효과를 낳았다. 여기서 더 나아가, 이렇게 축적된 다량의 생물기원 유기물이 생태계에 미칠 잠재적인 되먹임(feedback) 효과를 고려해 볼 수 있다. 비록 본 연구에서 직접 측정하지는 않았지만, 선행 연구들에 따르면 이러한 신선하고 분해되기 쉬운(labile) 유기물은 이종영양세균의 성장을 급격히 촉진하는 우수한 기질로 작용한다(Azam et al. 1983). 세균 군집의 활발한 분해 활동은 수층, 특히 성층이 발달한 저층의 용존산소를 빠르게 소모하여 저산소(hypoxia) 환경을 유발할 수 있으며, 이는 저서생물에게 치명적인 영향을 미칠 수 있다(Rabalais 2002). 또한, 유기물 분해 과정(재광물화, remineralization)을 통해 유기물에 결합되어 있던 질소와 인이 다시 무기 영양염 형태로 수층에 공급될 수 있다. 이는 또 다른 식물플랑크톤의 성장을 자극하여 적조를 장기화하거나 2차 대발생을 유발하는 ‘양성 되먹임 고리(positive feedback loop)’로 작용할 가능성을 내포한다(Heisler et al. 2008). 따라서 본 연구의 결과는 남해안의 요소 유입이 단순히 일회성 적조 발생으로 끝나는 사건이 아님을 시사한다. 이는 자생적 유기물 생성이라는 매개체를 통해, 생태계 전체의 구조와 기능을 장기간에 걸쳐 변화시킬 수 있는 강력한 교란 요인임을 의미한다.
4. 결 론
본 연구는 2023년 여름철, 질소가 제한된 한국 남해안에서 요소의 생지화학적 역할과 유해성 적조 대발생과의 연관성을 분석하였다. 연구 결과, 여름철 몬순 시기에 집중적으로 유입되는 요소는 DIN과 함께 식물플랑크톤 군집의 성장을 촉발하는 주요 동인(key driver)으로 작용하였다. 요소는 DON 풀 내에서 생체이용가능한 질소원이었으며, 그 공급 증가는 이 해역에서 매년 여름철 반복되는 Cochlodinium polykrikoides 적조 발생의 잠재적 위험을 높이는 시공간적 패턴과 일치하였다. PCA 분석은 적조 발생 과정이 7월의 영양염 유입에 의한 ‘유발 단계’와 8월의 고수온에 의한 ‘유지/심화 단계’로 구분될 수 있다는 모델을 뒷받침한다. 본 연구 결과는 그동안 국내 연안의 부영양화 관리 및 적조 연구에서 상대적으로 간과되어 온 유기질소, 특히 요소의 중요성을 부각시킨다. 향후 연안 환경의 수질 관리 및 적조 예보의 정확도를 향상시키기 위해서는 DIN 중심의 모니터링 체계에서 벗어나 요소를 포함한 생체이용가능 DON과 육상 오염원 관리를 포함하는 통합적 접근이 필요하다.
