

1. 서 론
2. 재료 및 방법
조사지 개황
현장조사
군집분석
건강도 평가
3. 결 과
출현 종수 및 개체수
우점종
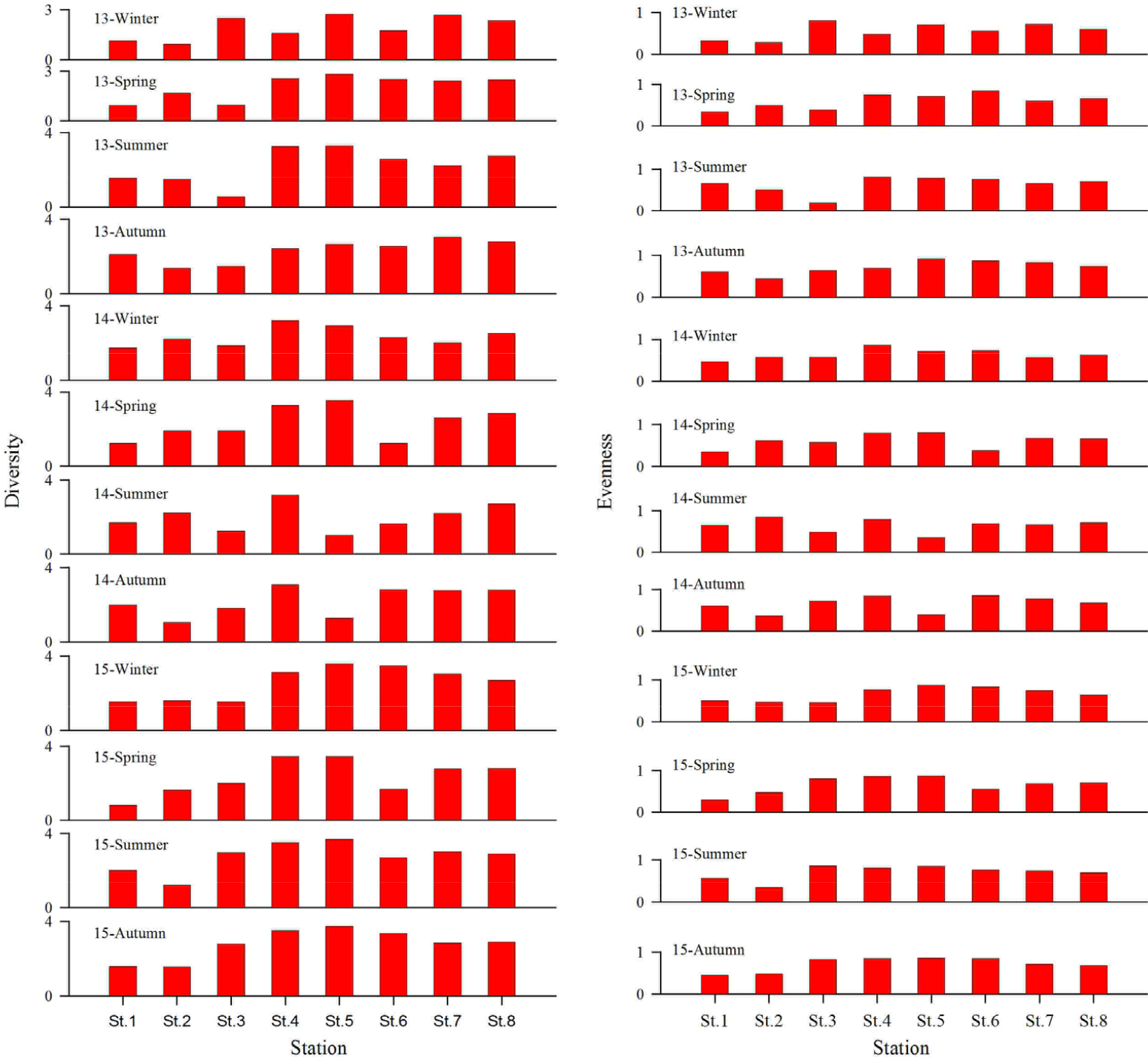
생태학적 제 지수 및 군집구조
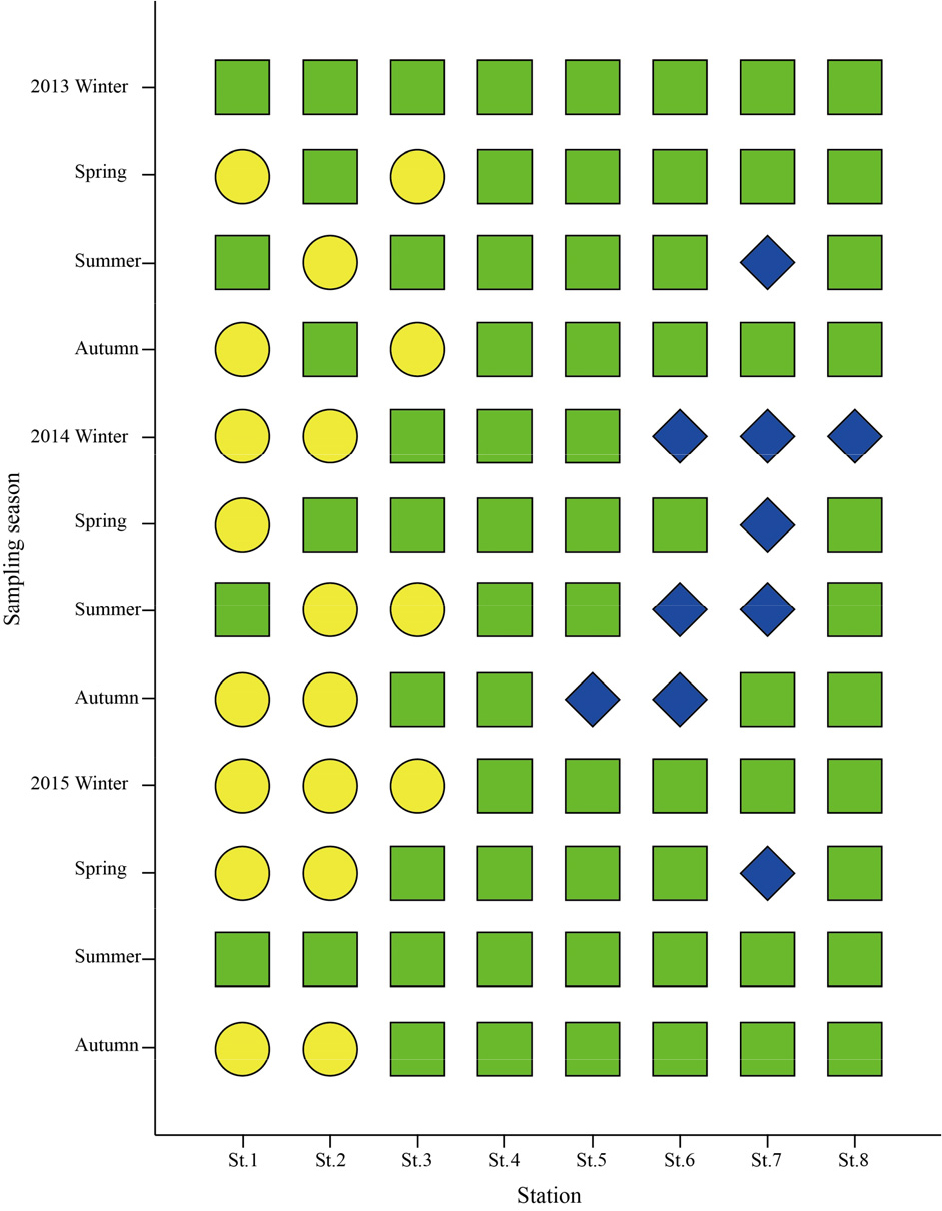
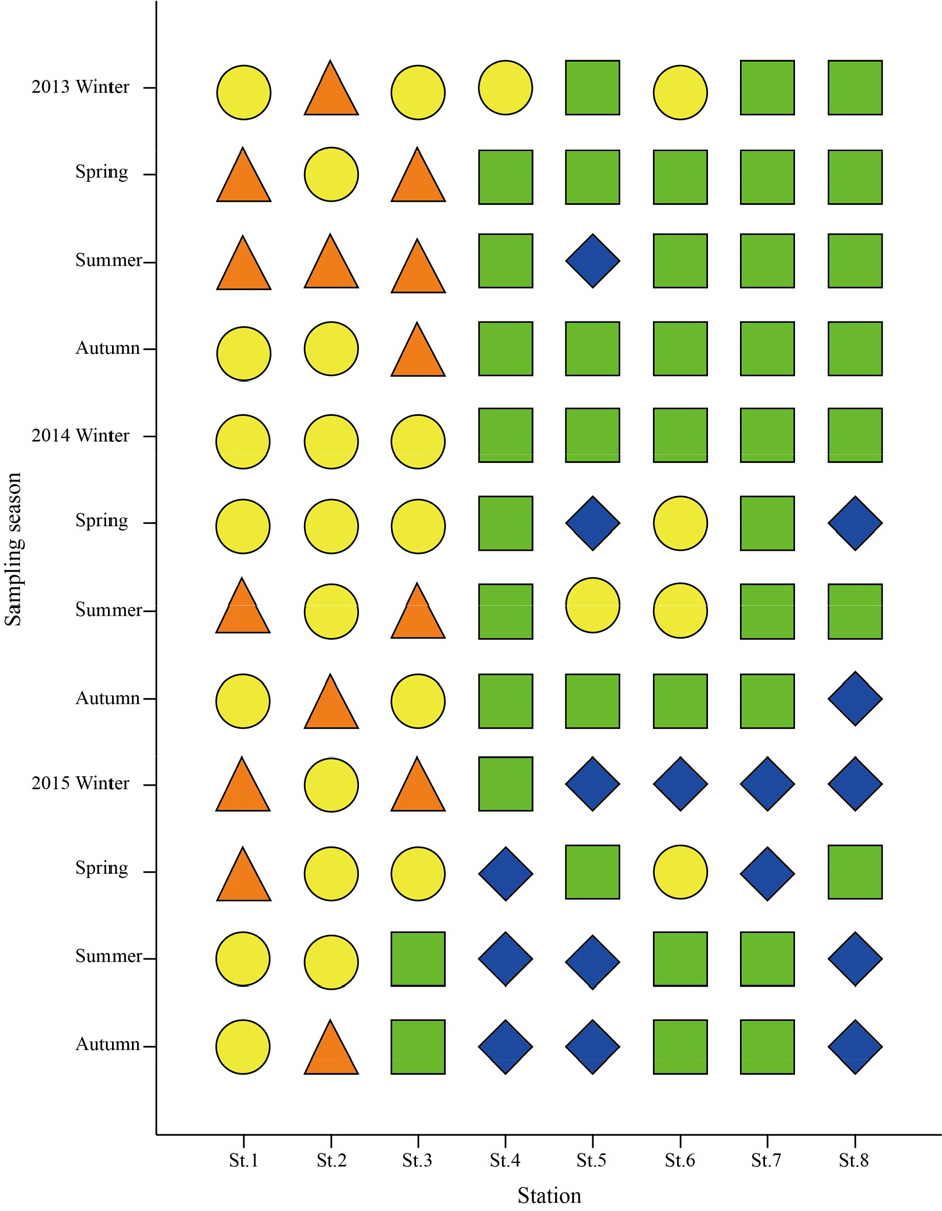
건강도 평가
4. 고 찰
대형저서동물의 생물량, 우점종 및 군집구조
건강도
5. 결 론
1. 서 론
하구역은 육지로부터 공급되는 담수와 해수의 혼합으로 인해 희석된 염분농도를 가지는 반폐쇄 형태의 연안수역이며(Pritchard 1967), 다양한 물질이 집적되고 유출되는 육상과 해양간의 여과장치로서 역할을 수행한다(Schubel and Kennedy 1984; McLusky 1989). 또한 하구역은 담수 유입량의 정도, 바람 및 조석 등 다양한 환경요인에 의해 수괴가 완전히 혼합되지 않는 이질적인 상태를 유지함으로서 수온, 염분농도와 용존산소농도 등 환경요인의 시・공간적인 변화가 매우 크다(Kikuchi 1981; Benyoucef et al. 2013). 따라서 하구역에 서식하는 대형저서동물은 환경적인 스트레스에 대한 적응내성이 뛰어남과 동시에 군집구조는 매우 복잡한 시․공간적인 변화양상을 나타낸다(Ysebaert et al. 2003). 이렇듯 하구역에 서식하는 대형저서동물 군집의 생물다양성은 인접한 해양 및 담수생태계와 비교해 상대적으로 낮지만, 환경에 적응한 특정 생물종의 개체군은 매우 잘 발달하여 단위면적당 생물량이 높다는 특징이 있다(Remane and Schlieper 1958; McLusky and Elliot 2004). 하구역을 포함한 연안역은 전 세계적으로 생물생산력이나 생태학적 가치가 매우 높은 생태계로 인식되고 있다(Day et al. 1989; Houde and Rutherford 1993; Costanza et al. 1997).
대형저서동물은 해양생태계를 구성하는 중요 구성생물군의 하나로 생물다양성의 가치가 매우 높다(van der Veer et al. 1990). 또한 대형저서동물은 이동성이 적거나 또는 정주성(고착성)의 특징을 보이며, 급성 또는 만성적인 서식처 교란에 반응하여 능동적으로 도피할 능력이 미약하기 때문에 환경변화의 영향을 감지하는데 있어 유용한 생물군이다(Pearson and Rosenberg 1978). 이렇듯 대형저서동물 군집은 서로 다른 형태의 환경교란에 적응내성을 가지는 다양한 생물군을 포함하고 있으며, 퇴적물의 내부 또는 표면 위를 흐르는 해수와의 영양염 순환(또는 물질순환)과 상위 영양단계로의 에너지 전달에 중요한 생태적인 역할을 담당하고 있다(Diaz et al. 2004; Dauvin et al. 2007). 결국 하구역에서 대형저서동물의 출현 종수, 개체수와 군집구조의 파악은 해역의 구조적 안정성과 환경교란의 정도를 감시하는데 있어 중요하다(Parker 1975; Rosenberg and Nilsson 2005; Wildsmith et al. 2009).
현재 해양생태계 연구에 있어서 대형저서동물을 대상으로 하는 다양한 생물 평가지수들이 개발되고 있고, 이러한 생물 평가지수들은 서로 다른 형태의 교란으로부터 생태계의 질적인 상태(quality status)를 평가하고 감시하는데 있어 효율적인 수단이 되고 있다(Borja et al. 2003). 특히 최근에는 정량적인 형태의 건강도 평가를 위하여 대형저서동물을 이용한 다양한 생물지수들이 지속적으로 개발 및 적용 중에 있다. 이 중 본 연구에 적용된 AMBI (ATZI’s Marine Biotic Index, Borja et al. 2003)와 M-AMBI 지수(Multivariate-AMBI, Muxica et al. 2007)는 전 세계적으로 무산소와 빈산소해역, 부영양화해역, 해사채취와 준설해역, 유류오염해역, 생활 및 산업오폐수 유출해역 등 인위적인 교란이 다양하게 수렴되는 환경에 대한 감지와 평가능력이 높은 것으로 입증되고 있다(Borja et al. 2009). 또한 본 연구에서는 낙동강 하구역 주변해역에 다수의 양식어장이 존재한다는 점에서 「어장환경평가의 방법 및 절차 등에 관한 규정(국립수산과학원 고시 제2014-08호 2014)」에서 제안하고 있는 BHI (어장관리법 시행규칙 제3조)지수를 기초로 저서생태계의 건강도를 비교함으로서 현 상태를 객관적으로 검증하고자 한다.
지금까지 낙동강 하구역 주변해역에서 수행된 대형저서동물 연구는 크게 군집과 생물상 연구로 양분되어 있다. 이 가운데 군집연구로는 Lee et al. (2005) 및 Seo et al. (2016)과 Bae and Yoon (1988, 1989a, 1989b)의 결과로 전자는 하구역의 외해역을 대상으로, 후자는 낙동강 하구역의 기수역 및 모래 등을 대상으로 수행하였다는 점에서 차이가 있다. 또한 Yoon et al. (2011)은 낙동강 하구역 전역을 대상으로 홍수기 전후에 따른 대형저서동물 군집의 변화상을 밝힌 바 있다. 반면 생물상 연구로는 주로 대형저서동물군의 대표적 생물군인 연체동물과 절지동물의 갑각류를 대상으로 실시된 바가 있다(Kim and Jang 1987; Kwon 1988; Jo 1988a, 1988b, 1989, 1990a, 1990b; Kim 1991; Jang and Kim 1992). 또한 낙동강 하구역의 건강도 평가는 해양역을 중심으로 수행된 연구가 유일하다(Seo 2016). 본 연구는 낙동강 하구역의 기수역과 해양역을 대상으로 대형저서동물 군집에 대해 3개년 간의 비교적 장기간의 현장조사를 기초로 대형저서동물 군집의 시·공간적인 변화특성과 건강도를 제시하는데 목적이 있다.
2. 재료 및 방법
조사지 개황
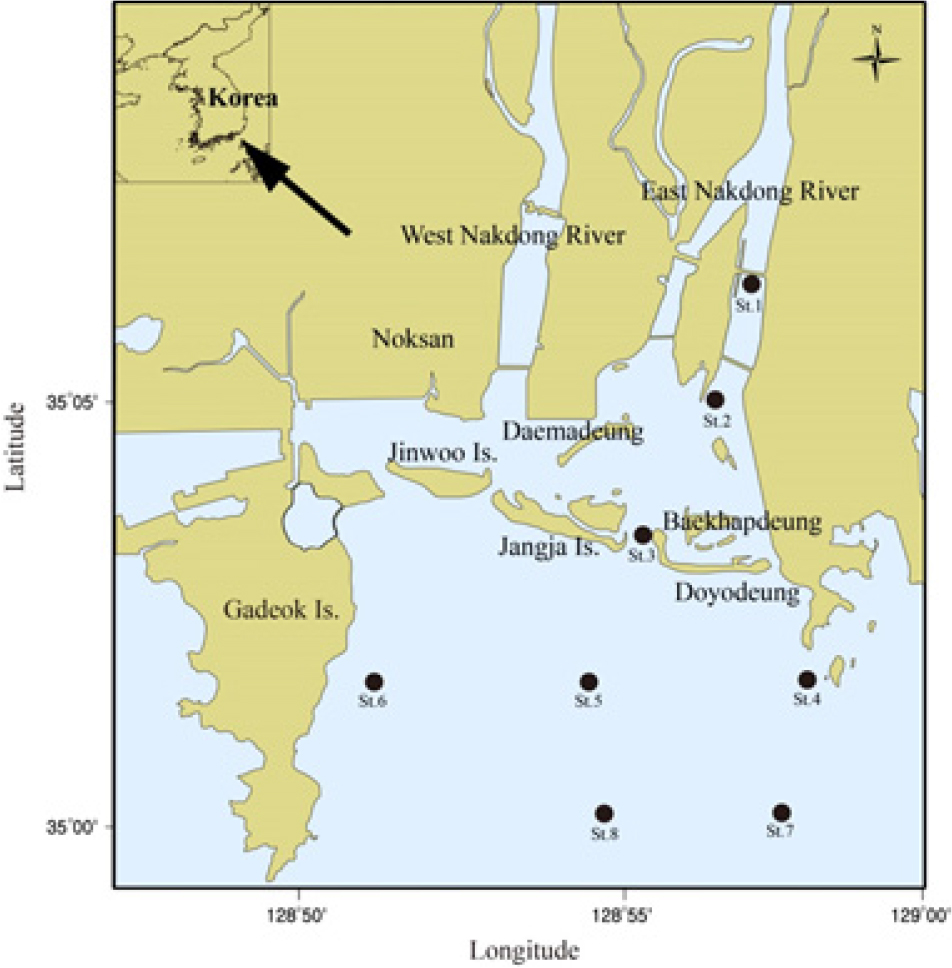
낙동강은 강원도 삼척시 황지에서 발원하여 총 연장 522 km를 흐르는 국내에서 2번째로 긴 하천이며, 23,656 km2에 달하는 집수역의 물을 모아 최종적으로 남해로 유입된다(Park et al. 2001). 또한 낙동강은 서해안으로 열려있는 강과 비교해 상대적으로 넓은 면적의 기수역을 가지고 있으며, 하구역에는 갯벌, 삼각주 및 모래등(진우도, 대마등, 장자도, 맹금머리등, 백합등, 도요등)과 같이 다양한 서식처가 존재한다. 이렇듯 낙동강 유역 일원해역은 1966년에 천연기념물 제179호(문화재청)로 지정된 것을 시작으로 시간경과에 따라 부산연안특별관리해역(1982), 자연환경보전지역(1987), 자연생태계보전지역(1989) 및 습지보호지역(1999)으로 지정․보호되고 있다. 반면 낙동강 유역 일원해역은 1960년대와 1990년대에 각각 산업단지와 녹산․명지지구 조성을 위한 간척사업이 실시된 바 있고, 특히 1983년부터 1987년까지 농업용수의 원활한 공급을 도모하고자 하굿둑 조성공사가 이루어졌다. 또한 1990년대 후반과 2000년대 중․후반에는 부산신항, 을숙도 대교 및 4대강 사업에 따른 상류지역의 보건설과 동시에 준설작업이 수행된 바 있다. 현재까지도 낙동강 유역 일원해역에서는 에코델타시티조성사업이 진행 중에 있어 보전과 개발압력이 상충하는 장소로 고려할 수 있다.
현장조사
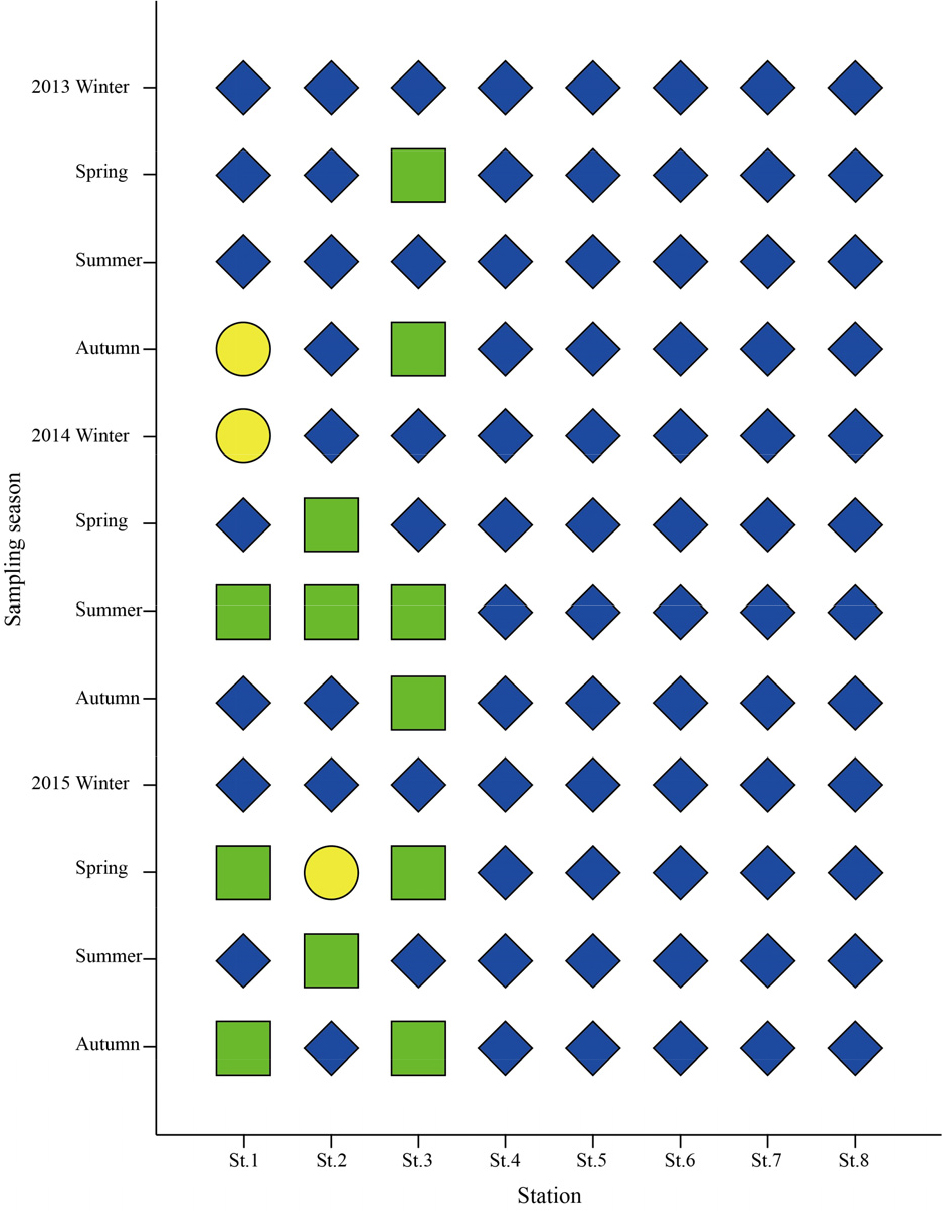
연구해역은 낙동강 하굿둑에서 외해역까지 총 8개 정점을 대상으로 하였고, 대형저서동물의 채집을 위한 현장조사는 2013년부터 2015년까지 계절별로 실시하였다(Fig. 1). 또한 현장조사 정점은 본 조사와 동시에 이루어진 Yoo et al. (2016)과 Yoon et al. (2017)의 염분농도 자료를 기초로 기수역 3개 정점(정점 1~3)과 외해역 5개 정점(정점 4~8)이 위치하고 있다. 한편 대형저서동물의 채집은 채집면적이 0.1 m2인 개량된 채니기(modified van Veen grab sampler)를 이용하여 정점 당 2회(0.2 m2)의 퇴적물을 채취하였다. 현장조사에서 채취된 퇴적물은 1 mm 망목의 체를 이용하여 대형저서동물을 일차적으로 분리하였고, 10%의 중성 포르말린으로 고정하여 실험실로 운반한 후 분류군별로 선별하였다. 한편 선별된 표본은 가능한 종 수준까지 동정을 실시하였고, 개체수를 계수하였다.
군집분석
연구해역에 서식하는 대형저서동물의 군집구조를 파악하기 위하여, 종 다양도 지수(H’, Shannon and Weaver 1949)와 균등도 지수(Pielou 1977)를 구하였다. 한편 출현 종과 개체수 자료를 이용한 군집분석에서 각 정점 간의 유사도(similarity)를 파악하기 위하여, Bray and Curtis (1957)의 지수를 이용하였다. 유사도 지수 행렬로부터 각 조사 정점과 출현 종을 연결하는 방법으로는 group-average를 이용하였다. 군집분석은 PRIMER (Plymouth Routines Multivariate Ecological Research) computer package를 이용하여 수지도(Dendrogram)로 표현하였다(Clarke and Warwick 2001). 또한 연구해역에서 출현한 대형저서동물의 시·공간에 따른 출현 종수와 개체수의 차이유무를 파악하기 위하여 일원일차분산분석(one-way ANOVA)을 실시하였다. 일원일차분산분석에 앞서 분산의 동질성 파악을 위해 레빈검정(Levene’s test)을 수행하였고, 원자료는 필요한 경우에 로그변환하였다. 연구해역의 우점종은 출현 개체수를 기초로 선정하였다.
건강도 평가
연구해역에 서식하는 대형저서동물의 개체수 자료를 기초로 건강도를 평가하였다. 건강도 평가는 유럽연합에서 이용되고 있는 AMBI (AZTI’s Marine Biotic Index, Borja et al. 2003) 및 M-AMBI지수(Multivariate-AMBI, Muxica et al. 2007)와 국내에서 개발된 BHI지수를 산출하여 비교를 시도하였다. 이 중 AMBI지수는 개별 종의 민감도와 내성도(sensitivity and tolerance)를 파악하여 해당 기능군의 점유율을 기초로 생산되며, 그 평가는 AZTI’s Marine Institute에서 제공하는 소프트웨어(www.azti.es)를 이용하여 산출하였다. 반면 BHI지수는 환형동물문에 속하는 다모류(해산갯지렁이류) 개별종의 유기물 농도에 따른 출현 빈도와 밀도를 기초로 분류된 4개의 기능군에 대한 개체수를 기초로 산출하였다. 특히 본 지수의 이용에 있어서 개별 기능군의 분류는 다모류 그룹별 종 목록(국립수산과학원 고시 2014-08호)을 활용하였다.
3. 결 과
출현 종수 및 개체수
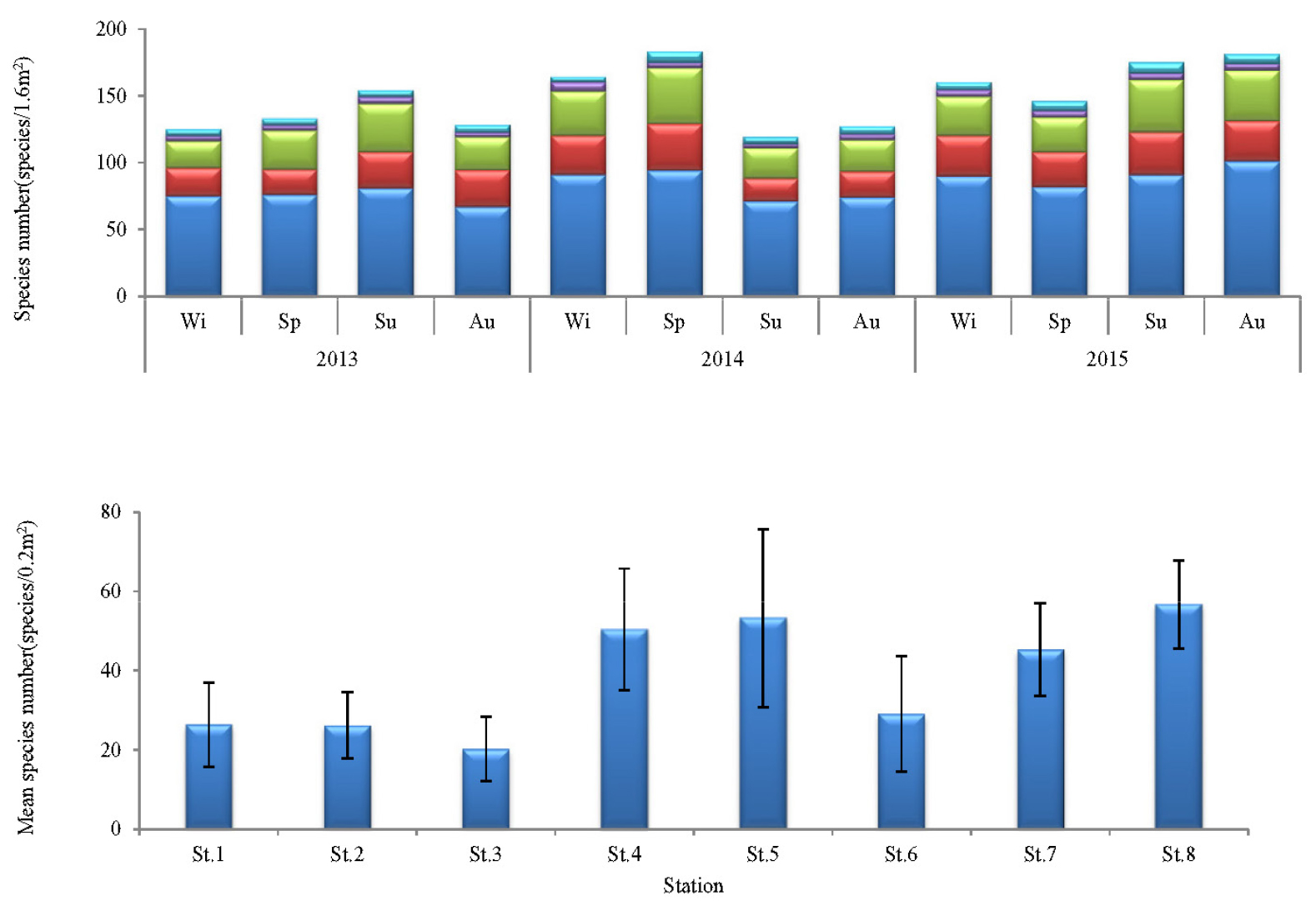
연구해역에서 출현한 대형저서동물은 총 380종/19.2 m2이었고, 출현 개체수는 441,855개체(4,603개체/m2)를 나타내었다. 출현 종수는 환형동물의 다모류가 152종(40.3%)이 채집되어 가장 우점하였고, 다음으로 절지동물의 갑각류와 연체동물이 각각 108종(28.7%)과 98종(25.3%)이 출현하였다(Fig. 2). 한편 조사시기에 따른 출현 종수는 119~ 183종의 범위에 2014년 하계에 가장 적었고, 동년 춘계에 가장 많았다. 이 밖에 2013년 하계, 2014년 동계, 2015년 동계, 하계 및 추계에 150종 이상이 채집되어 상대적으로 다양하였다. 또한 조사정점별 평균 출현 종수는 20~57종의 범위에 정점 3에서 가장 적었고, 정점 8에서 가장 많았다. 이 외에 정점 4, 5와 7에서 각각 평균 50종, 53종 및 45종이 출현하여 상대적으로 많았다(Figs. 3 and 5).
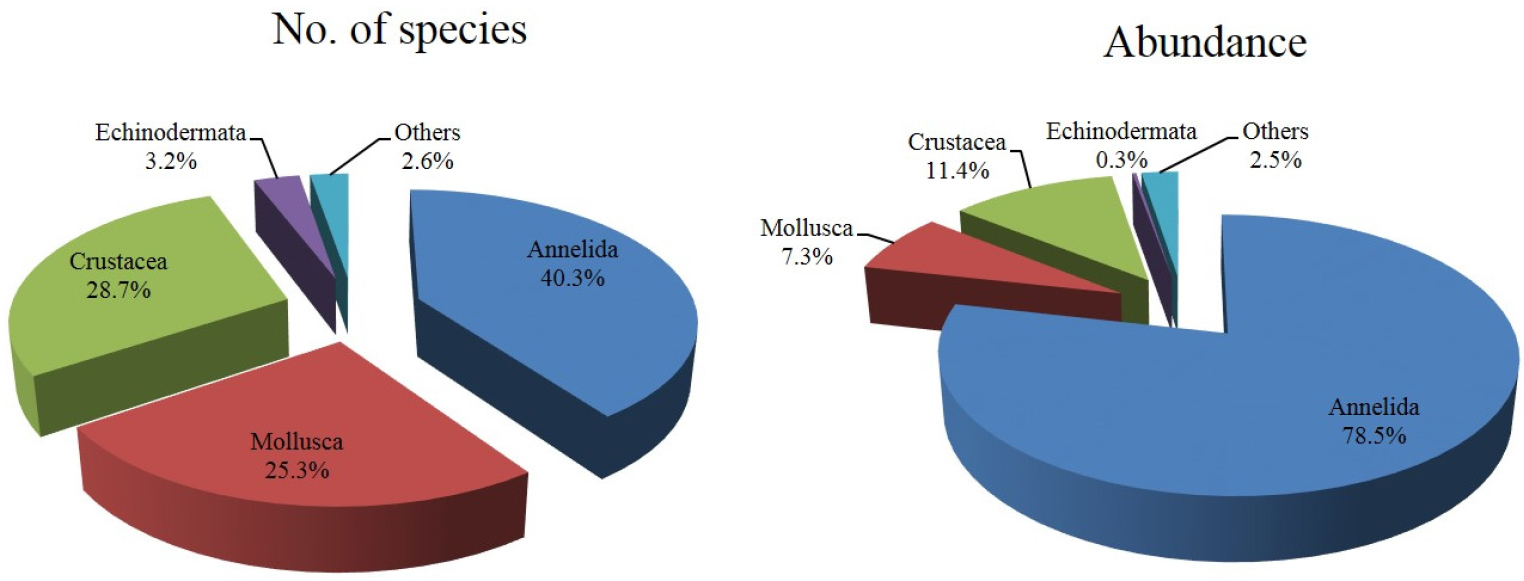
출현 개체수에 있어서도 다모류는 346,900개체(78.5%)로 가장 많았고, 다음으로 갑각류, 연체동물 및 기타동물의 순서로 각각 50,310개체(11.4%), 32,315개체(7.3%) 및 11,115개체(2.5%)가 채집되었다(Fig. 2). 출현 개체수는 조사시기에 따라 7,705~74,065개체의 범위에 2013년 추계에 가장 적었고, 동년 동계에 가장 많았다. 이 밖에 2014년 동계, 2015년 동계 및 하계에 40,000개체 이상의 밀도를 나타내 상대적으로 많았다. 또한 조사정점별 평균 출현 개체수는 904~19,722개체의 범위에 정점 6에서 가장 적었고, 정점 1에서 가장 많았다. 이 외에 정점 2에서 7,844개체가 출현하여 상대적으로 많았다. 이상을 종합하면, 연구해역에서 출현한 대형저서동물의 출현 종수는 시간경과에 따라서 증가의 경향을 나타낸 반면, 개체수는 감소와 증가의 경향을 보여 차이가 있었다. 반면 공간적으로도 출현 종수는 해양역 정점군에서 많았고, 출현 밀도는 기수역 정점군에서 상대적으로 높은 값을 보여 차이가 있었다(Figs. 4 and 5).
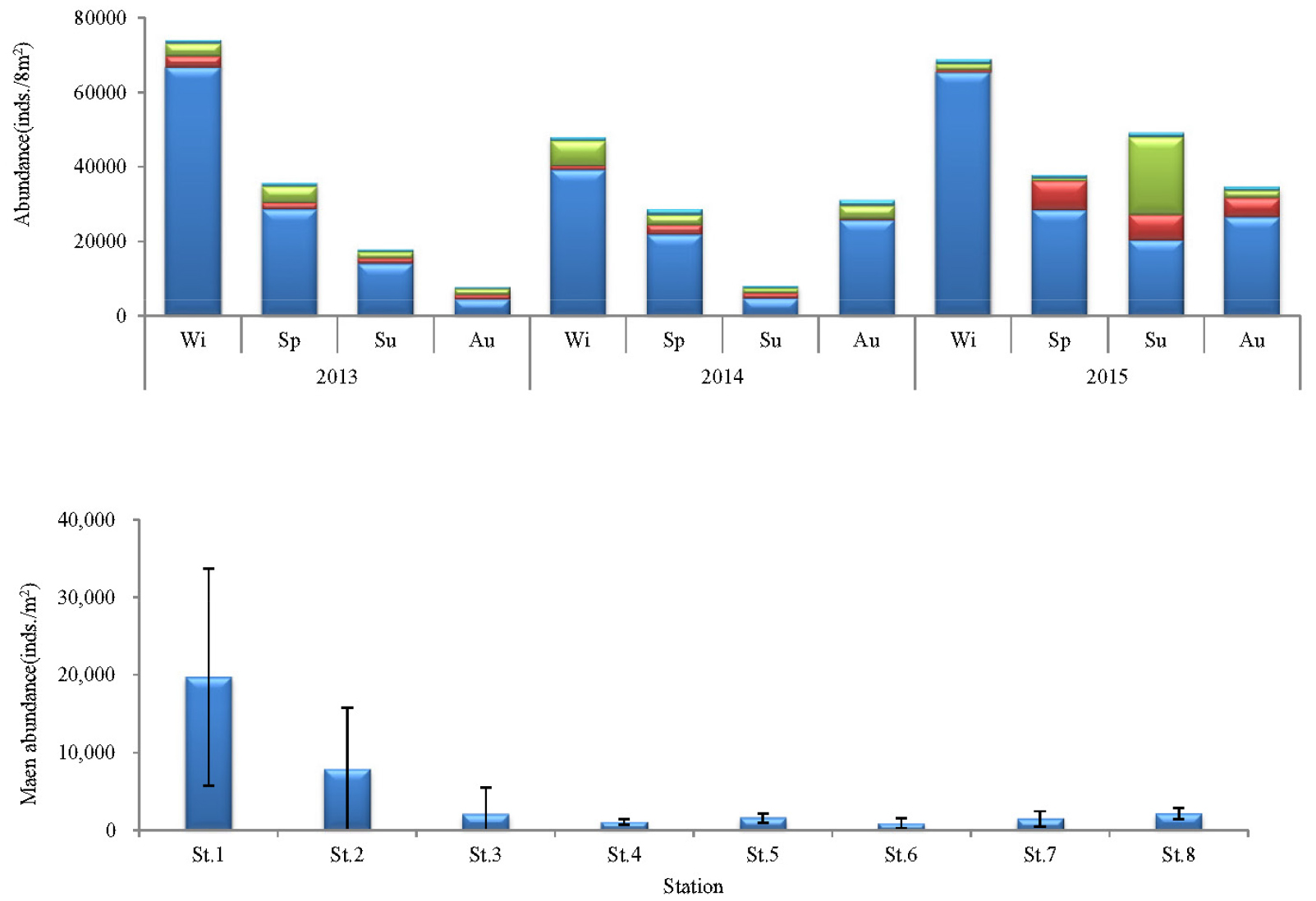
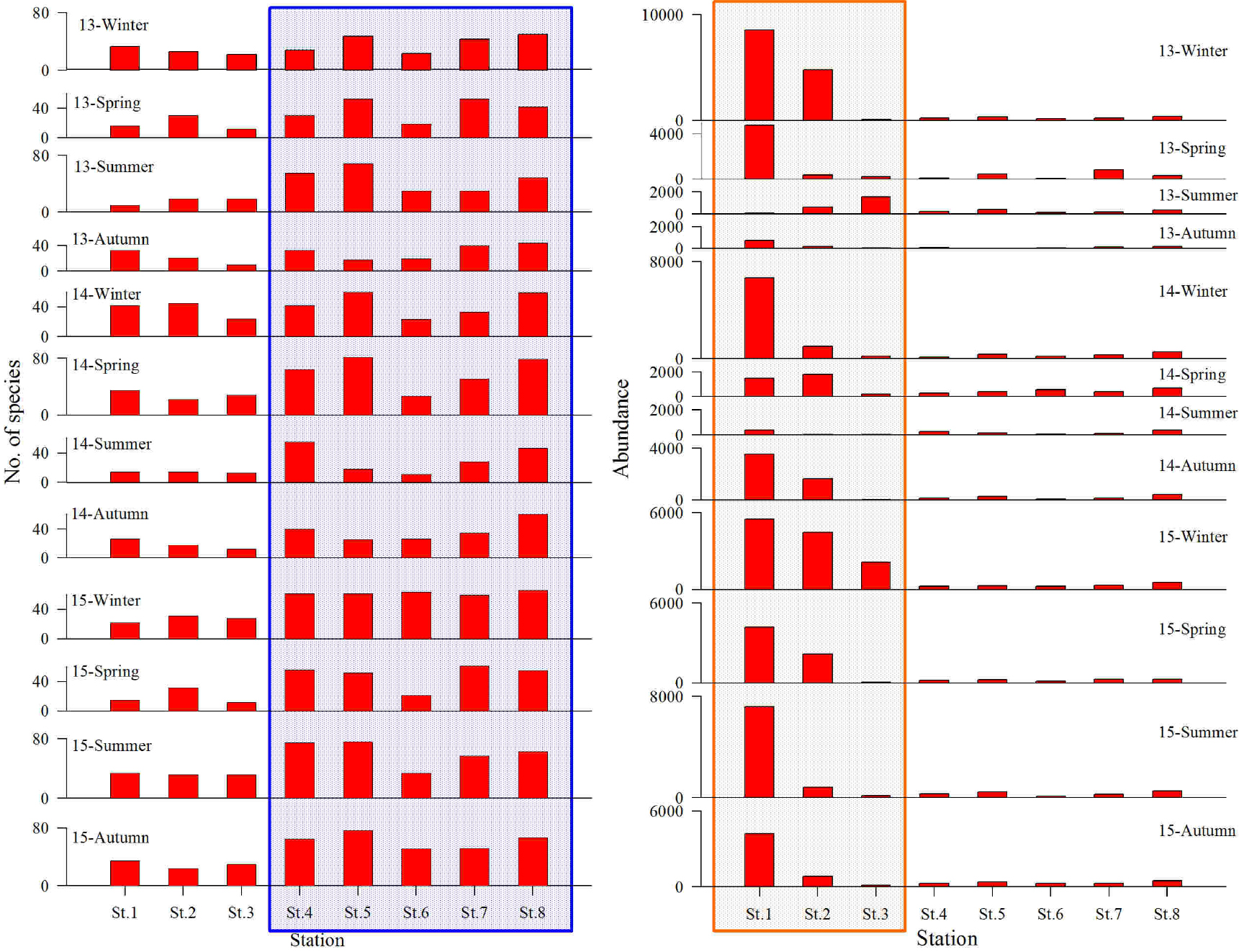
한편 대형저서동물의 출현 종수와 개체수의 조사시기 및 정점에 따른 차이유무를 파악하기 위하여 실시한 일원일차분산분석에서도 평균 출현 종수는 차이가 있었고, 특히 2015년에 평균 출현 종수가 많았다(F2, 95 = 5.03, p < 0.01). 또한 조사정점에 따른 평균 출현 종수(F7, 95 = 13.109, p < 0.01)와 개체수(F7, 95 = 12.620, p < 0.01)는 해양역과 기수역 정점군에서 차이가 있었고, 출현 종수는 해양역의 정점군에서 많았던 반면, 개체수는 기수역의 정점군에서 많았다.
우점종
연구해역에서 출현한 대형저서동물의 개체수를 기준으로 상위 10위 우점종을 선정하였다. 분류군에 있어서는 다모류가 8종으로 가장 많았고, 다음으로 갑각류와 연체동물의 이매패류가 공통적으로 1종이 상위에 위치하였다. 이들을 최우선 순위별로 보면, 다모류의 Minuspio japonica, 두점박이선녀얼굴갯지렁이(Pseudopolydora kempi), 고리버들갯지렁이(Heteromastus filiformis), 갑각류의 발성육질꼬리옆새우(Grandidierella japonica), 다모류의 Polydora cornuta, 등가시버들갯지렁이(Capitella capitata), 연체동물 이매패류의 종밋(Arcuatula senhousia), 다모류의 양손갯지렁이(Magelona japonica), 나팔얼굴갯지렁이(Rhynchospio glutaea) 및 작은부채발갯지렁이(Eteone longa) 등이었다. 이들 상위 10위 우점종들은 총 321,130개체의 밀도를 나타내 전체의 72.7%를 차지하였다. 특히 Minuspio japonica는 88,205개체가 출현하여 전체의 20.0%로 가장 점유율이 높았고, 다음으로 두점박이선녀얼굴갯지렁이가 79,620개체, 18.0%를 점유하였다(Table 1).
Table 1.
Top 10 dominant species ranked by their total abundance in the Nakdong River Estuary, Busan, Korea during this study period (Apol: Annelida Polychaeta, CAm: Arthropoda Crustacea Amphipoda, MBi: Mollusca Bivalvia)
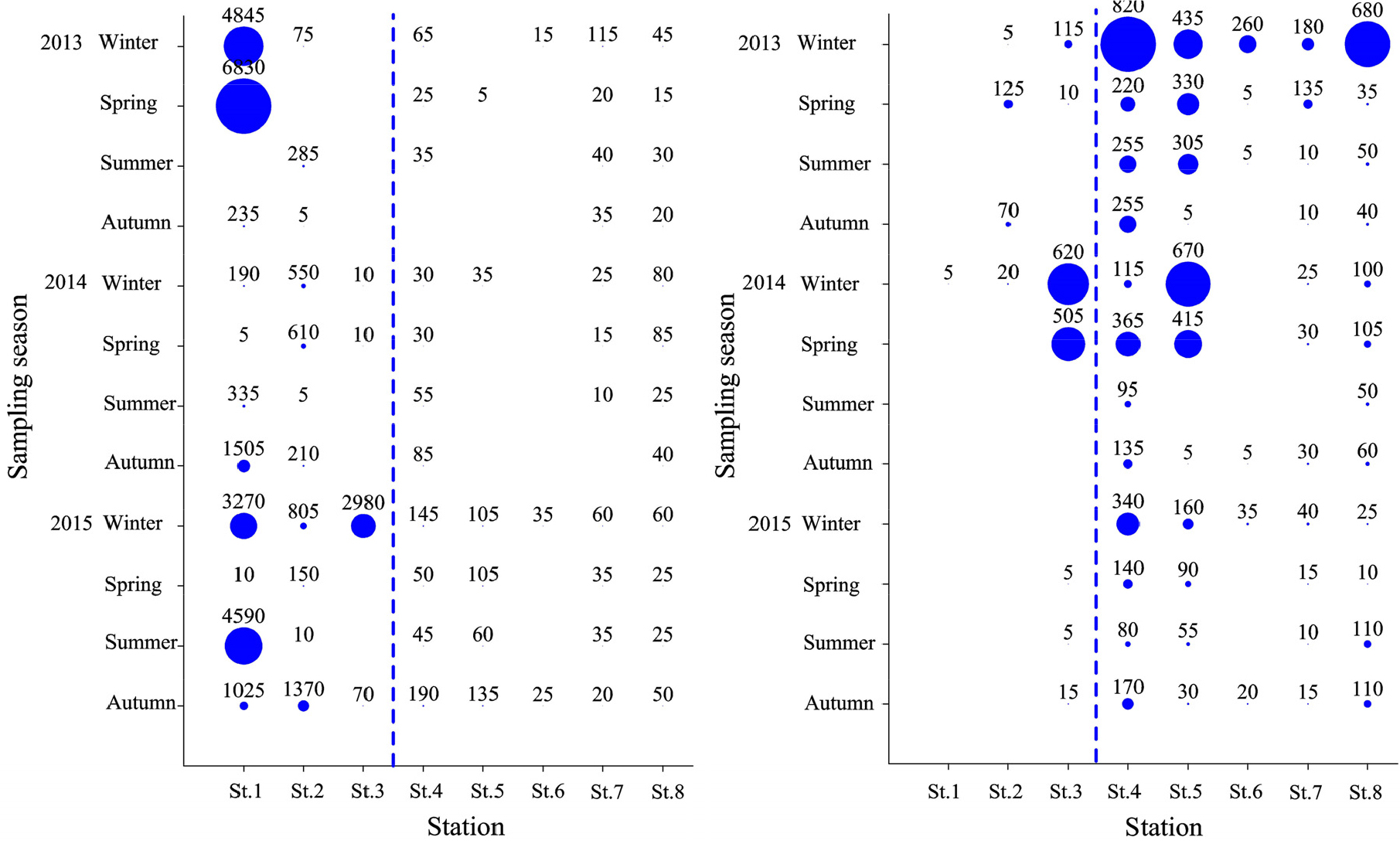
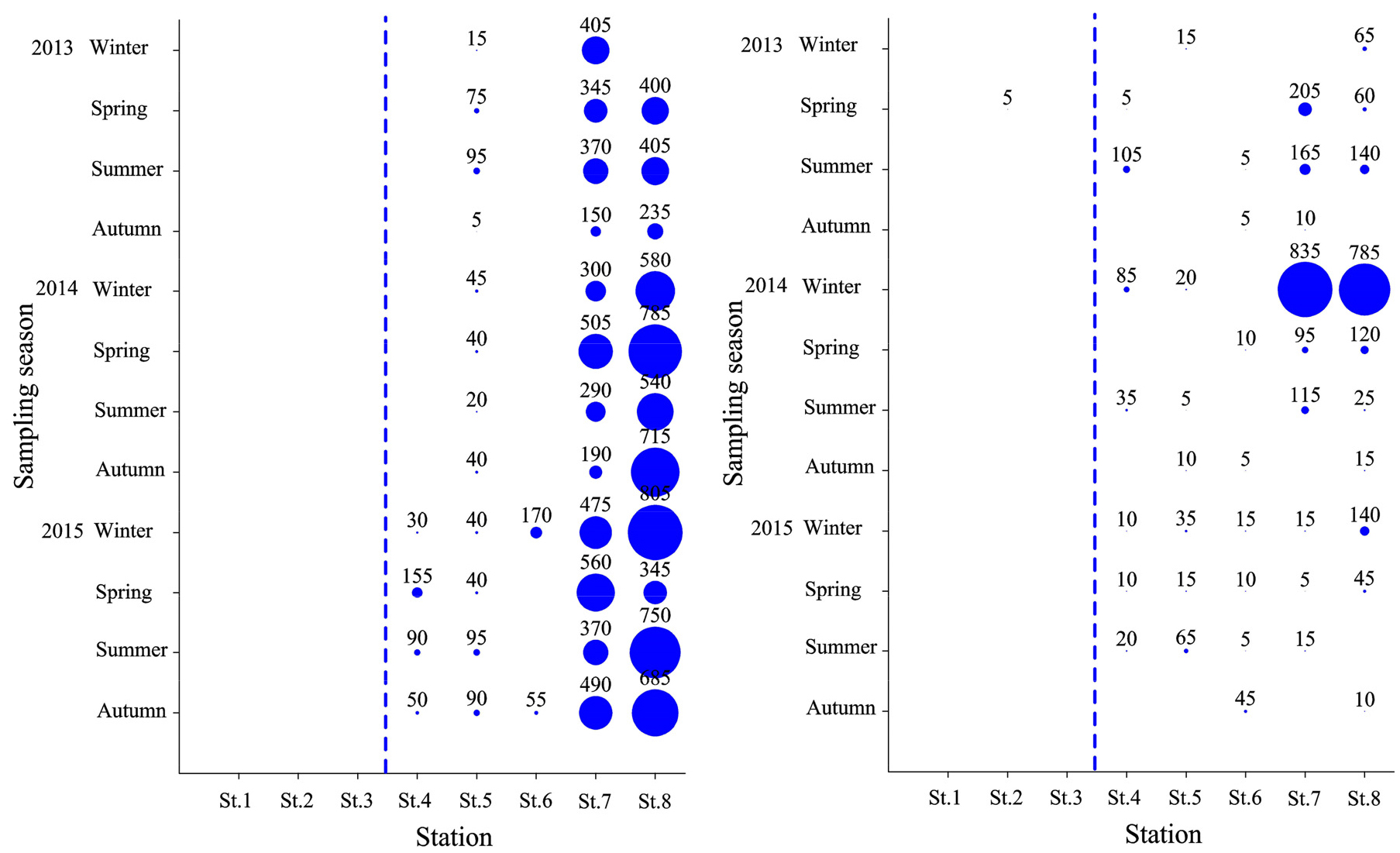
이상에서와 같이 연구해역에서 출현한 대형저서동물의 개체수 기준 상위 우점종은 공간적으로 광역분포종, 해양역 분포종 및 기수역 분포종으로 크게 구분할 수 있었다. 이 중 광역분포종은 다모류의 고리버들갯지렁이와 오뚜기갯지렁이가 대표적이었다. 그러나 전자는 공간분포가 기수역에 보다 집중되었고, 조사시기에 따라서도 2013년 동계(5,160개체), 춘계(6,895개체), 2015년 동계(7,460개체) 및 하계(4,765개체)에 특징적으로 밀도가 높았다. 반면 후자는 해양역에 보다 넓게 분포하고 있었고, 조사시기별로는 2013년 동계(2,495개체), 2014년 동계(1,555개체)와 춘계(1,420개체)에 밀도가 높았다(Fig. 6). 다음으로 해양역 분포종은 양손갯지렁이와 갑각류의 안경옆새우류인 Ampelisca bocki가 이에 해당하였다. 이 가운데 양손갯지렁이는 해양역의 모든 정점에서 출현하고 있었으나, 특히 정점 7과 8에서 그 밀도가 높았다. 그러나 본 종의 조사시기에 따른 밀도 변화는 상대적으로 크지 않았다. 또한 A. bocki도 해양역의 전 정점에서 채집되었고, 조사시기별로는 2014년 동계에 정점 7과 8에서 각각 835개체와 785개체로 많았던 것을 제외하면, 기타 조사시기에는 비교적 밀도가 적었다(Fig. 7). 마지막으로 기수역 분포종은 Minuspio japonica, 두점박이선녀얼굴갯지렁이, 등가시버들갯지렁이, 발성육질꼬리옆새우 및 종밋 등이었다. 이들 5종은 기수역의 정점 1, 2 및 3에서 공통적으로 밀도가 높았다. 이 중 Minuspio japonica는 2013년 하계와 2015년 동계 및 춘계에 각각 16,365개체, 26,000개체 및 17,455개체가 채집되어 비교적 춘계와 조사후반기인 2015년에 전반적으로 밀도가 높았다. 다음으로 P. kempi는 2013년과 2015년 동계에 각각 50,025개체와 15,820개체가, 등가시버들갯지렁이는 2014년 동계에 15,640개체가 채집되어 출현 밀도가 매우 높았다. 한편 발성육질꼬리옆새우는 2015년 하계에 14,320개체가 출현하여 특징적이었고, 종밋은 2015년 춘계부터 추계에 이르기까지 각각 6,205개체, 5,045개체 및 4,425개체가 출현하였다(Fig. 8).
생태학적 제 지수 및 군집구조
연구해역에서 대형저서동물 군집의 다양도 지수는 0.55~3.73의 범위에 2013년 하계의 정점 3에서 가장 낮았고, 2015년 추계의 정점 5에서 가장 높았다. 다양도 지수는 기수역의 정점 1, 2 및 3에서 전반적으로 낮은 값을 나타낸 반면, 해양역의 정점군에서 상대적으로 높은 값을 보였다. 그러나 해양역 정점군에서는 2014년 하계에 다양도 지수 값의 감소가 관찰되었다(Fig. 9). 한편 균등도 지수는 0.19~0.91의 범위에 2013년 하계의 정점 3에서 가장 낮았고, 동년 추계의 정점 5에서 가장 높았다. 이렇듯 2013년 하계의 낮은 균등도 지수는 다모류의 Minuspio japonica와 두점박이선녀얼굴갯지렁이의 대량출현에 그 원인이 있었다(Fig. 9).
연구해역에서 출현한 대형저서동물의 개체수를 기초로 실시한 집괴분석 결과에서 대형저서동물 군집은 크게 3개의 그룹으로 구분되었다. 이 중 그룹 1은 기수역에 위치한 정점 1, 2 및 3으로 구성되었고, 다모류의 Minuspio japonica, 두점박이선녀얼굴갯지렁이 및 고리버들갯지렁이가 우점하는 군집이었다. 다음으로 그룹 2는 주로 해양역의 정점 4, 5, 7 및 8을 포함한 가운데 다모류의 양손갯지렁이, 오뚜기갯지렁이 및 성구동물의 상어껍질별벌레(Phascolosoma scolops)가 우점하였다. 마지막으로 그룹 3은 가덕도 인근의 정점 6을 대표하였고, 다모류의 큰갈매기고리갯지렁이(Goniada japonica), Mediomastus californiensis 및 옆새우류의 Eochelidium miraculum이 우점하는 군집이었다(Fig. 10; Table 2). 한편 상기의 3개 군집 중에서 출현 종수는 그룹 2에서 평균 51종이 출현하여 가장 많았고, 다음으로 그룹 1과 3에서 공통적으로 24종이 채집되었다. 반면 개체밀도는 그룹 1에서 평균 11,710개체로 가장 높았고, 그룹 2와 3에서 각각 평균 1,525개체와 805개체가 출현하였다. 다양도 지수는 그룹 2에서 평균 2.85의 수치로 가장 높았고, 다음으로 그룹 3과 1에서 각각 평균 2.13과 1.50을 나타내었다. 따라서 연구해역의 대형저서동물 군집은 해양역의 정점군에서는 출현 종수와 다양도 지수가 높았던 반면, 기수역의 정점군에서는 특정 종의 대량 출현에 의해 개체수가 많다는 차이를 보였다.
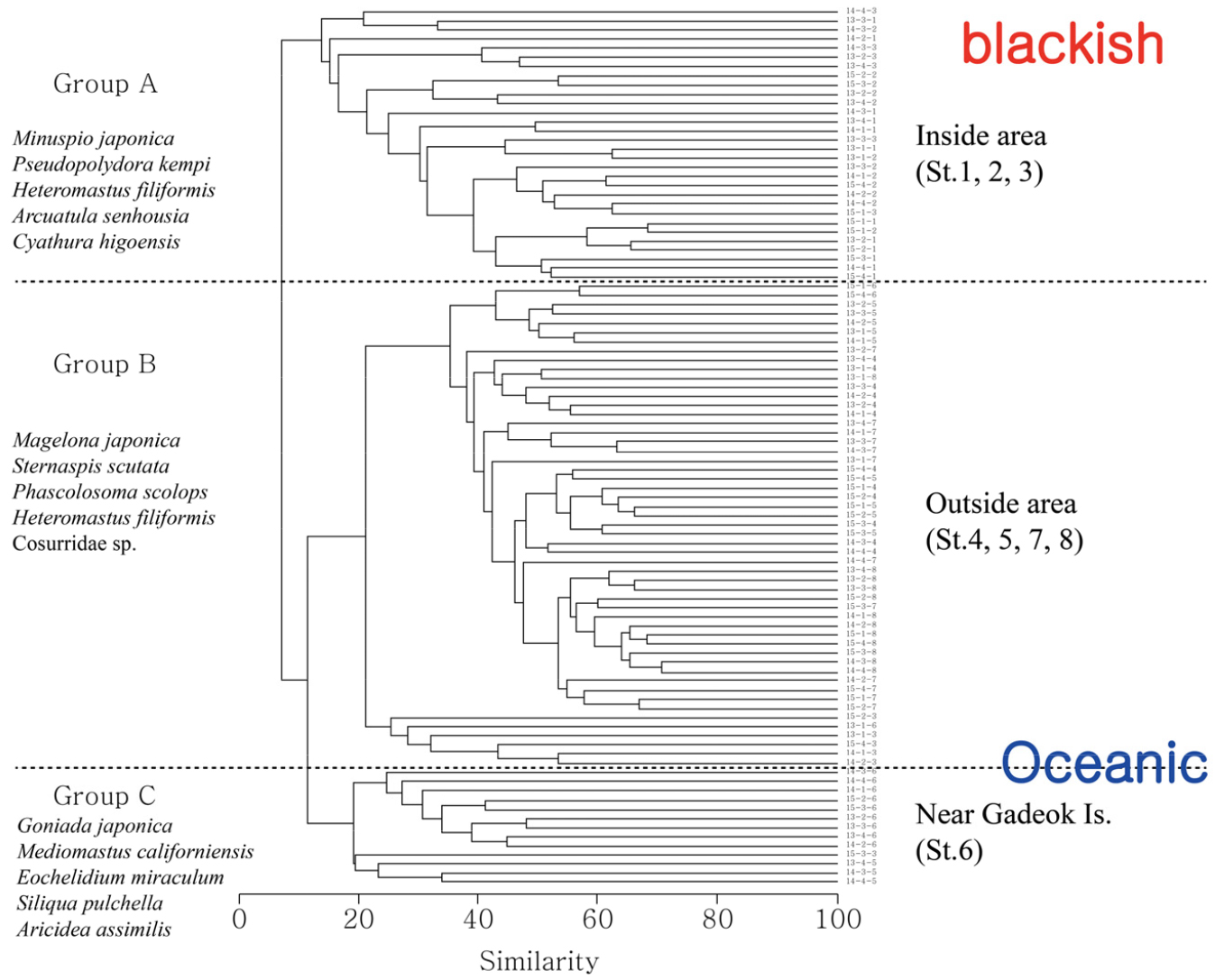
Table 2.
Ecological characteristics of each station group determined by cluster analysis in the Nakdong River Estuary, Busan, Korea
건강도 평가
AMBI
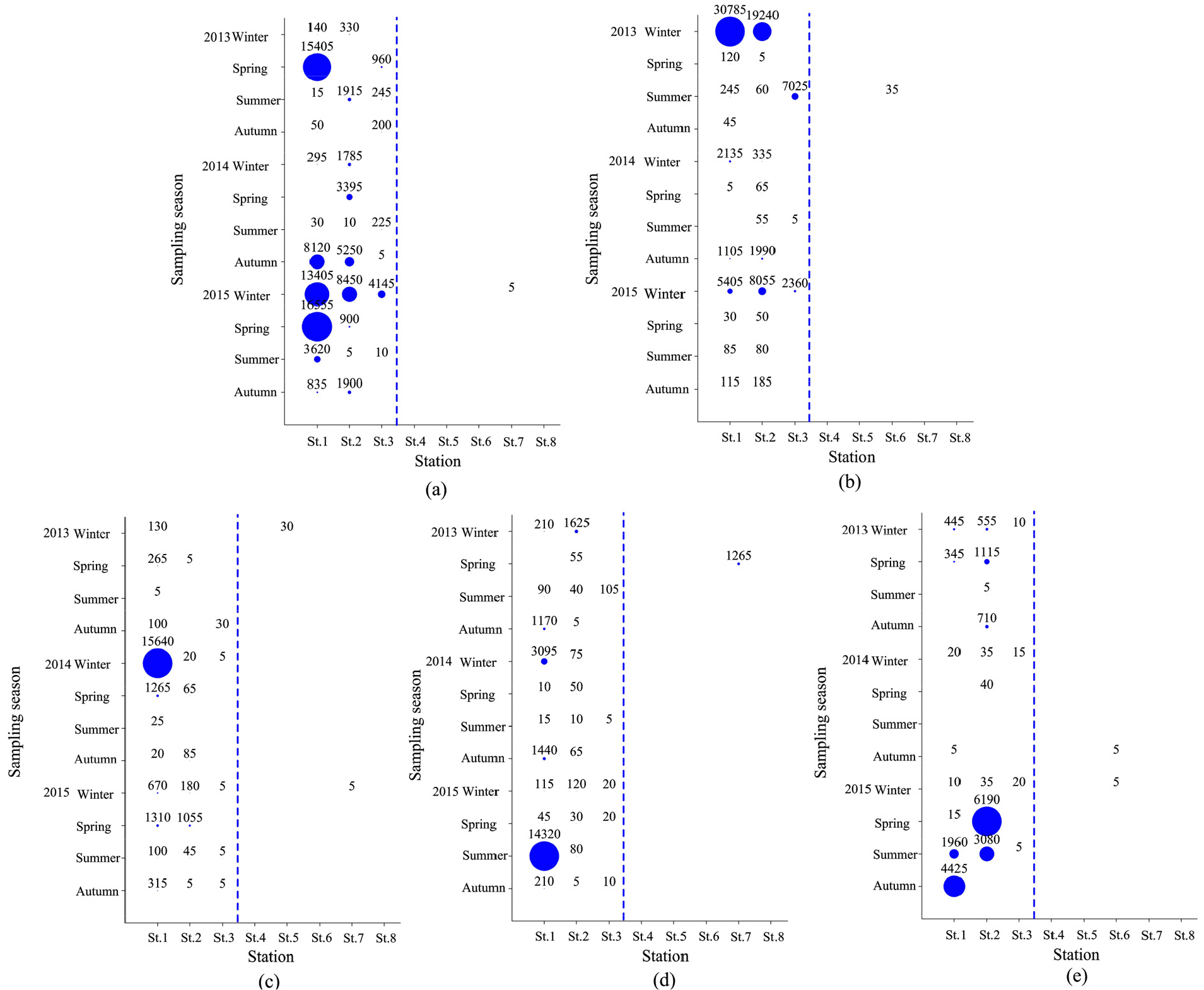
연구해역에서 AMBI지수는 0.4~4.7의 범위에 2014년 추계의 정점 5에서 가장 낮았고, 동년 동계의 정점 1에서 가장 높았다. 연구해역의 총 96개 정점을 대상으로 한 AMBI지수에서 정상 또는 교란되지 않은 상태(undisturbed status)는 2014년 추계의 정점 5를 포함한 총 10개 정점으로 구성되었고, 다음으로 약간 교란 상태(slightly status)는 2013년 동계의 정점 1을 비롯한 총 67개 정점이, 마지막으로 중간 교란 상태(moderately status)는 2014년 동계의 정점 1을 포함한 총 19개 정점이었다. 반면 심한 교란 상태(Extremely disturbed status)를 나타내는 정점은 없었다. AMBI지수를 기초로 한 연구해역의 건강도는 전반적으로 기수역의 정점군에서 교란 정도가 비교적 심하였던 반면, 해양역의 정점군에서 상대적으로 양호하였다(Fig. 11).
M-AMBI
연구해역에서 M-AMBI지수는 0.23~0.89의 범위에 2015년 춘계의 정점 1에서 가장 낮았고, 2014년 춘계의 정점 5에서 가장 높았다. 연구해역의 총 96개 정점을 대상으로 한 M-AMBI지수에서 가장 양호한 상태(high status)는 2014년 춘계의 정점 5를 포함한 총 16개 정점이 포함되었고, 다음으로 양호한 상태(good status)와 중간 상태(moderate status)는 각각 40개(2013년 동계의 정점 5 등)와 26개 정점(2013년 동계의 정점 1 등)이었다. 반면 불량한 상태(poor status)를 나타내는 정점은 2015년 춘계의 정점 1을 비롯한 총 14개 정점이 포함되었고, 가장 불량한 상태(bad status)의 정점은 없었다. M-AMBI지수를 기초로 한 연구해역의 건강도는 전반적으로 기수역의 정점군에서 중간에서 불량한 상태를 반영하는 정점이 많았던 반면, 해양역의 정점군에서는 비교적 양호 또는 가장 양호한 상태를 나타낸 정점들로 구성되었다(Fig. 12).
BHI
연구해역에서 BHI지수는 32~98의 범위에 2014년 동계의 정점 1에서 가장 낮았고, 2013년 하계의 정점 3에서 가장 높았다. 연구해역의 총 96개 정점을 대상으로 한 BHI지수에서 가장 양호한 상태(1등급)는 2013년 하계의 정점 3을 포함하여 총 81개 정점으로 구성되었고, 다음으로 양호한 상태(2등급)는 2013년 춘계의 정점 3을 비롯하여 총 12개 정점이었다. 반면 불량한 상태(3등급)는 2013년 추계의 정점 1을 포함한 3개 정점이, 그리고 가장 불량한 상태(4등급)의 정점은 없었다. BHI지수를 기초로 한 연구해역의 건강도는 2013년 춘계와 2014년 동계의 정점 1과 2015년 하계의 정점 2를 제외하면 전반적으로 모든 정점에서 2등급 이상의 양호한 환경상태를 반영하고 있었다(Fig. 13).
4. 고 찰
낙동강 하구역의 주변해역은 습지 및 사주가 잘 발달한 전형적인 하구역의 공간적 특성을 가지나, 인근해역에는 대도시와 다수의 공단 및 항만시설이 입지하는 특수성이 있다. 또한 연구해역은 1987년 11월에 하굿둑이 완공되면서 담수와 해수의 자연적인 혼합이 인위적으로 통제되는 등 자연 하천의 기능이 차단되어 생태계 변화가 비교적 심한 환경으로 분류할 수 있다(Yoon et al. 2011). 한편 우리나라 서해안에 위치한 한강, 금강, 만경강 및 동진강 하구역의 대형저서동물 군집은 일차적으로 염분농도의 구배에 의해, 또한 공간분포 양상은 퇴적상에 의해 영향을 받는 것으로 보고된 바 있다(Choi and Koh 1994; Yoo and Hong 1996). 본 연구에서는 대형저서동물의 서식에 영향을 미치는 환경요인에 대한 측정은 이루어지지 않았다. 그러나 본 연구와 동일한 시기에 측정된 Yoon et al. (2017)에 의하면, 연구해역은 하굿둑 건설 이전에는 담수유입과 조석작용에 의해 일반적인 하구역의 해수유동을 보이나 하굿둑 완공이후에는 담수 유입량에 의해 조절되는 양상으로 변경되었고, 하굿둑에서 해양역으로의 염분농도 구배가 현재까지도 비교적 뚜렷한 것으로 제시되고 있다. 또한 서식처의 기질로서 퇴적상에 있어서도 현장조사시의 관찰과 기존 연구(Yun and Paik 2001; Lee et al. 2005; Yoon et al. 2011; Seo et al. 2016)를 종합해 보면, 낙동강 하굿둑에서 해양역까지는 전반적으로 펄퇴적물이 우세하게 분포하나, 가덕도 인근해역은 세립한 모래퇴적물로 구성되었음을 알 수 있다.
대형저서동물의 생물량, 우점종 및 군집구조
연구해역에서 출현한 대형저서동물은 총 380종에 단위 면적당(m2) 출현 개체수는 4,603개체이었다. 본 연구에서의 380종은 가덕도 주변해역의 260종(Yun and Paik 2001), 가덕도 주변과 해양역의 203종(Lee et al. 2005), 기수역과 해양역의 150종(Yoon et al. 2011) 및 해양역의 266종(Seo et al. 2016)등과 비교해 상대적으로 생물다양성이 높다. 또한 단위면적당(m2) 출현 개체수에 있어서도 Yun and Paik (2001)의 1,729개체, Lee et al. (2005)의 1,554개체와 Seo et al. (2016)의 859개체보다도 월등히 높은 밀도를 나타내고 있다. 이상에서와 같이 상기 연구들은 공간적으로 낙동강 하구일원 해역을 대상으로 하고 있다는 점에서 공통점이 있다. 그러나 Lee et al. (2005)과 Yoon et al. (2011)은 각각 2001년과 2010년에 실시된 일회성 조사라는 점에 차이가 있다. 반면 Seo et al. (2016)은 본 연구와 동일하게 3개년의 조사와 계절성이라는 조사주기를 반영하고 있다는 점에서 공통점이 있으나, 공간적으로는 해양역과 기수역-해양역의 일부 정점을 대상으로 한다는 점에서 차이가 있다. 이렇듯 동일한 해역을 대상으로 하는 연구에서 현장조사 시의 채집면적과 빈도(횟수)등의 조사강도와 서식처의 퇴적상은 대형저서동물의 출현 종수와 밀도 차이에 영향을 미친다. 따라서 본 연구에서는 채집면적이 서로 다른 해역에서의 출현 종수 비교를 위하여 이용되는 Whittaker (1975)의 지수를 도입하여 비교를 시도하였다(Table 3). 그 결과, 본 연구에서 296으로 가장 높았고, 다음으로 241 (Yun and Paik 2001), 220(Lee et al. 2005), 207(Seo et al, 2016) 및 193(Yoon et al. 2011)의 수치를 보여 본 연구의 지수가 기존 연구결과와 비교해 높은 값을 보이고 있어 생물다양성이 높다는 것을 간접적으로 입증하고 있다. 결국 본 연구에서의 높은 출현 종수와 밀도는 상기의 연구들이 단기적인 연구기간(일회성과 계절성)과 특정 해역(해양역 및 기수-해양역)을 대상으로 한다는 점에서 발생하는 차이로 고려할 수 있다. 한편 연구해역에서 대형저서동물의 출현 종수는 시간경과에 따라 지속적으로 증가하는 경향을 보였고, 특히 2014년 이후에는 동계에 공통적으로 출현 종수가 많았다. 반면 개체수는 감소와 증가의 경향을 반복적으로 나타내고 있으나 출현 종수의 경향과 유사하게 매년 동계에 공통적으로 밀도가 높았다. 한편 공간적으로 출현 종수는 해양역의 정점군에서 상대적으로 다양하였던 반면, 개체수는 공통적으로 기수역의 정점군에서 많았다. 또한 다양도와 균등도 지수도 생물지수의 경향을 잘 반영하여 기수역 정점군에서는 낮은 값을, 반대로 해양역 정정군에서는 높은 값을 나타내었다. 일반적으로 자연상태의 하구역에 서식하는 대형저서동물의 종 다양성은 담수종과 해산종의 임계 염분농도(critical salinity)인 5–8 psu에서 최소가 되는 것으로 알려져 있다. 공간적으로도 강을 향해서는 담수종이, 해양을 향해서는 해산종이 증가하게 된다(Khlebovich 1968; Remane and Schlieper 1971). 텍사스의 Corpus Christi Bay 하구역에서 조사한 결과를 보면, 출현 종수는 담수가 유입되는 곳으로부터 해양으로 향하면서 증가하는 양상을 보이는 반면, 밀도는 감소하는 것으로 알려져 있다(Flint and Kalke 1985). 이러한 출현 종수와 개체수의 분포 양상은 본 연구해역에서도 잘 일치하고 있다.
Table 3.
Sampling area, species numbers and abundances of the present and previous studies in the Nakdong River Estuary, Busan, Korea (Whittaker’s d is also presented for the comparison of relative species diversity)
| Sampling area (m2) | Species number | Abundance (ind./m2) | Whittaker d | Reference |
| 12 | 260 | 1,729 | 241 | Yun and Paik 2001 |
| 8.4 | 203 | 1,554 | 220 | Lee et al. 2005 |
| 6 | 150 | - | 193 | Yoon et al. 2011 |
| 19.2 | 266 | 859 | 207 | Seo et al. 2016 |
| 19.2 | 380 | 4,603 | 296 | Present study |
연구해역에서 출현한 대형저서동물의 주요 우점종은 공간적으로 광역분포종, 해양역 분포종 및 기수역 분포종으로 크게 구분할 수 있다. 이 중 광역분포종은 다모류의 고리버들갯지렁이와 오뚜기갯지렁이가 대표적이었다. 그러나 전자는 그 분포가 기수역에 보다 집중되어 있고, 후자는 해양역에 넓게 분포하고 있다. 반면 해양역 분포종은 양손갯지렁이와 갑각류의 안경옆새우류인 Ampelisca bocki가 이에 해당하고, 기수역 분포종은 Minuspio japonica, 두점박이선녀얼굴갯지렁이, 등가시버들갯지렁이, 발성육질꼬리옆새우 및 종밋 등이었다. 이상에서 연구해역에 서식하는 주요 우점종은 염분농도의 구배에 따라 그 서식분포를 달리함을 알 수 있다. 이와 같은 결과는 대형저서동물 군집의 조사정점에 따른 차이유무를 파악하기 위한 통계분석에서도 잘 반영되어 평균 출현 종수와 개체수는 해양역과 기수역 정점군에서 유의한 차이가 있었고, 특히 출현 종수는 해양역의 정점군에서 많았던 반면, 개체수는 기수역의 정점군에서 많아 이를 입증하고 있다. 또한 연구해역에서 대형저서동물의 군집도 크게 3개의 그룹으로 구분된 가운데 그룹 1은 기수역에 위치한 정점 1, 2 및 3이, 그룹 2는 해양역의 정점 4, 5, 7 및 8로 구성되고 있다. 반면 그룹 3은 가덕도 인근의 모래퇴적물로 구성된 정점 6을 대표하고 있다. 결국 연구해역에 서식하는 대형저서동물 군집은 염분농도의 구배와 퇴적상의 차이에 따라 생물지수(출현 종수와 개체수), 우점종의 공간분포 및 구조가 결정되고 있음을 알 수 있다.
건강도
연구해역에 서식하는 대형저서동물 군집의 건강도는 3개 지수(AMBI, M-AMBI 및 BHI)에서 공통적으로 기수역의 정점군에서 불량(또는 교란 상태)한 환경상태를 나타낸 반면, 해양역의 정점군에서 양호한 환경상태를 반영하고 있다. 특히 불량한 환경상태를 나타내고 있는 기수역의 총 24개 정점들 가운데 AMBI와 M-AMBI지수는 교란 또는 불량한 상태를 반영하는 정점이 각각 19개와 14개로 많았으나, BHI지수는 불과 3개로 적게 나타나고 있다. 따라서 3개의 지수를 종합적으로 고려해 보면, AMBI와 M-AMBI지수 결과는 BHI지수와 비교적 차이를 보이고 있었고, 서식처의 환경상태는 BHI지수와 비교해 2개 지수가 보다 적절하게 표현하고 있는 것으로 판단된다. 따라서 BHI지수는 지수 개발의 목적과 부합되게 어류양식어장에 한정하여 적용되어야 함을 지적한다고 볼 수 있다. 한편 연구해역의 기수역 정점군에서 출현한 우점종들은 환경이 불량하거나 또는 교란된 해역에서 출현하는 종들을 다수 포함하고 있다. 이들 종은 Pearson and Rosenberg (1978)가 제시하고 있는 유기물 오염과 관련하여 기회종적 특성을 보이거나 또는 오염지표종으로 분류되는 종들이 대부분으로 다모류의 Minuspio japonica, 두점박이선녀얼굴갯지렁이, 등가시버들갯지렁이, 고리버들갯지렁이 및 이매패류의 종밋 등이 이에 해당한다. Park et al. (2016)은 하굿둑 완공 직후 기수역에 서식하는 갑각류가 사라지고, 연체동물의 출현 종수와 밀도도 급격하게 감소하며 다모류의 Prionospio membranacea, 두점박이선녀얼굴갯지렁이와 단각류의 중국뱀옆새우(Sinocorophium sinensis)와 같은 대표적인 유기물 오염지시종이 출현하여 하굿둑 완공이후 저서 환경이 점진적으로 오염되고 있음을 보고한 바 있다. 또한 Seo (2016)는 기수역의 건강도가 해양역보다는 대체적으로 교란의 정도가 높은 것으로 보고하고 있어 본 연구와 잘 일치하고 있다. 이상에서와 같이 본 연구에서 적용된 3개의 건강도 지수와 다양도 지수 등을 종합적으로 고려한 결과, 연구해역은 전반적으로 해양역보다 기수역의 건강도가 상대적으로 불량한 것으로 판단된다. 또한 건강도 지수중에서 생물지수와 우점종의 생태학적 특징을 전반적으로 고려할 때 M-AMBI지수가 다른 지수와 비교해 해역의 건강도를 보다 잘 반영하는 것으로 판단된다.
5. 결 론
본 연구는 낙동강 하구역에 서식하는 대형저서동물 군집과 저서생태계의 건강도를 평가하기 위하여 2013년 2월부터 2015년 10월까지 각 8개 정점을 대상으로 현장조사를 실시하였다. 연구해역에서 조사기간 동안 출현한 대형저서동물은 총 380종으로 단위면적당(m2) 출현 개체수는 4,603개체이었다. 연구해역에서 출현한 주요 우점종은 환형동물 다모류의 Minuspio japonica, 두점박이선녀얼굴갯지렁이(Pseudopolydora kempi), 고리버들갯지렁이(Heteromastus filifomis), 등가시버들갯지렁이(Capitella capitata), 절지동물 갑각류의 발성육질꼬리옆새우(Grandidierella japonica)와 연체동물 이매패류의 종밋(Arcuatula senhousia) 등이었다. 연구해역에서 관찰된 다모류의 일부 종은 서식처의 환경상태가 불량할 때 높은 밀도를 나타내는 기회종들로 구성되었다. 한편 연구해역에 서식하는 대형저서동물 군집은 염분 농도의 구배에 따라 기수역과 해양역 정점군으로 구분되었다. 또한 기수역 정점군은 출현 개체수가 많았던 반면, 해양역 정점군은 출현 종수와 다양도 지수가 상대적으로 높아 차이가 있었다. 연구해역에서 출현한 대형저서동물의 생태학적 특성과 개체수를 기초로 한 건강도 평가에서 3개 지수(AMBI, M-AMBI, BHI)는 공통적으로 해양역 정점군에서는 양호한 상태를 나타낸 반면, 기수역 정점군에서는 상대적으로 불량한 상태를 반영하였다. 또한 건강도 평가의 3개 지수중에서 M-AMBI지수는 다른 2개 지수들과 비교했을 때 연구해역 대형저서군집의 건강상태를 적절하게 반영하는 것으로 판단되었다.
