

1. Introduction
2. Materials and Methods
Sample collection and isolation
Morphological identification
Molecular and phylogenetic analysis
Determination of optimal culture conditions
Analyses for fatty acid composition of lipids
3. Results and Discussion
Morphological characteristics
Molecular identification and sequence analysis
Verification of the optimal cultivation conditions of the isolated strain
Fatty acid composition of lipids
1. Introduction
Microalgae are single-celled photosynthetic organisms that play a vital role in global biogeochemical cycles, particularly in carbon and nitrogen cycling, contributing to the stability of ecosystems and the health of aquatic food webs (Arrigo 2005; Jeong et al. 2010). Among them, green algae (Chlorophyta) demonstrate remarkable ecological flexibility, adapting to a wide range of habitats from freshwater to marine and even terrestrial environments (John et al. 2002; Tragin and Vaulot 2018; Aigner et al. 2020; Hyung et al. 2021; Kang et al. 2022). Beyond their role as primary producers (Falkowski and Raven 2007), green algae are valued as potential resources in various industries, such as biofuels, nutraceuticals, and pharmaceuticals, owing to their bioactive compounds, including lipids, essential fatty acids, and pigments (Raja et al. 2008; Wilson et al. 2020; Kang et al. 2024).
Despite the ecological and biotechnological importance of green algae, the taxonomy and phylogenetic understanding of these organisms remain incomplete (Champenois et al. 2015; Borowitzka 2016). Challenges in morphological differentiation, combined with limited DNA sequencing, result in frequent misidentifications that can impact ecological assessments and hinder biotechnological applications (Gantar and Svircev 2008; Champenois et al. 2015; Vardaka et al. 2016). Consequently, reliable molecular and morphological analyses of strains from diverse environments are essential. The isolation and detailed characterization of individual strains are crucial for accurately classifying species and fully realizing their commercial potential. Furthermore, the implementation of the Nagoya Protocol has highlighted the strategic importance of securing biological resources, such as microalgae, for national competitiveness and biological resource sovereignty, further underscoring the value of these resources (Avilés-Polanco et al. 2019; Heinrich et al. 2020).
In this context, the genus Tetraselmis, first established by F. von Stein in 1878 and comprising 35 accepted species, 1 variety, and 1 forma (Guiry and Guiry 2024), is notable for its adaptability across a range of aquatic environments, including marine, brackish, and freshwater systems (John et al. 2002; Mata et al. 2023), and for its extensive biotechnological potential, supporting applications in fields such as biofuels (Lim et al. 2015), aquaculture (Patrinou et al. 2022), and bioremediation (Segovia-Campos et al. 2024).
Tetraselmis marina (Cienkowski) Norris et al. (1980), a coastal green microalga within the genus Tetraselmis (Chlorodendraceae), is widely recognized for its adaptability and productivity, which make it a valuable candidate for various biotechnological applications (Mata et al. 2023). This microalga exhibits unique physiological traits, including its ability to produce lipids and carotenoids (Moussa et al. 2017), tolerate heavy metals (Cameron et al. 2018), and synthesize bioactive compounds (Trentin et al. 2022), all of which underscore its industrial significance. However, research on indigenous T. marina strains from Korea is lacking, even though these strains are likely to possess unique adaptations to local environments. The indigenous strains must first be isolated in pure culture and then characterized taxonomically, genetically, physiologically, and biochemically to maximize their potential.
Thus, this study aimed to explore the taxonomic, physiological, and biochemical characteristics of the indigenous T. marina MP0060 isolated from seawater off the Dokdo Islands, Korea. The specific objectives were to (1) comprehensively analyze the morphological and molecular characteristics of MP0060; (2) determine its optimal growth conditions under varying temperatures and light intensities; and (3) analyze its biochemical attributes, specifically focusing on fatty acid composition. The findings of this study are expected to enhance the taxonomic and physiological understanding of T. marina MP0060 and facilitate its broad industrial application, particularly in the fields of biofuels and essential fatty acids.
2. Materials and Methods
Sample collection and isolation
Plankton samples from the Dokdo Islands in the East Sea of Korea (37°14'25.86"N 131°51'57.72"E) were collected using a water sampler (Mezzanine Water Sampler; DAIHAN CHEMLAB, Seoul, Republic of Korea) in April 2016, when the water temperature was 17°C and the salinity was 34 practical salinity units (psu) (Fig. 1, Table 1). The samples were gently filtered through a 154 µm Nitex mesh (Nitex; Dynamic Aqua Supply Ltd., Surrey, BC, Canada) and placed in six-well tissue culture plates (Falcon 3046; Becton Dickinson, Lincoln Park, NJ, USA). A clonal culture of T. marina was established by using two serial single-cell isolations. Polycarbonate (PC) bottles containing F/2 medium (AlgaBoost TM F/2 2000x; AusAqua, Wallaroo, SA, Australia) and isolated T. marina cells were filled with filtered seawater, capped, and then incubated at 22°C under cool white fluorescent light (~60 µmol photons m-2 s-1, photosynthetically active radiation; PAR) with a 14:10 h light/dark cycle. As the concentration of T. marina was increased, the cells were transferred to 50, 125, and 500 mL PC bottles (Nalgene ClearBoy; Thermo Fisher Scientific, Rochester, NY, USA) containing fresh F/2 medium.
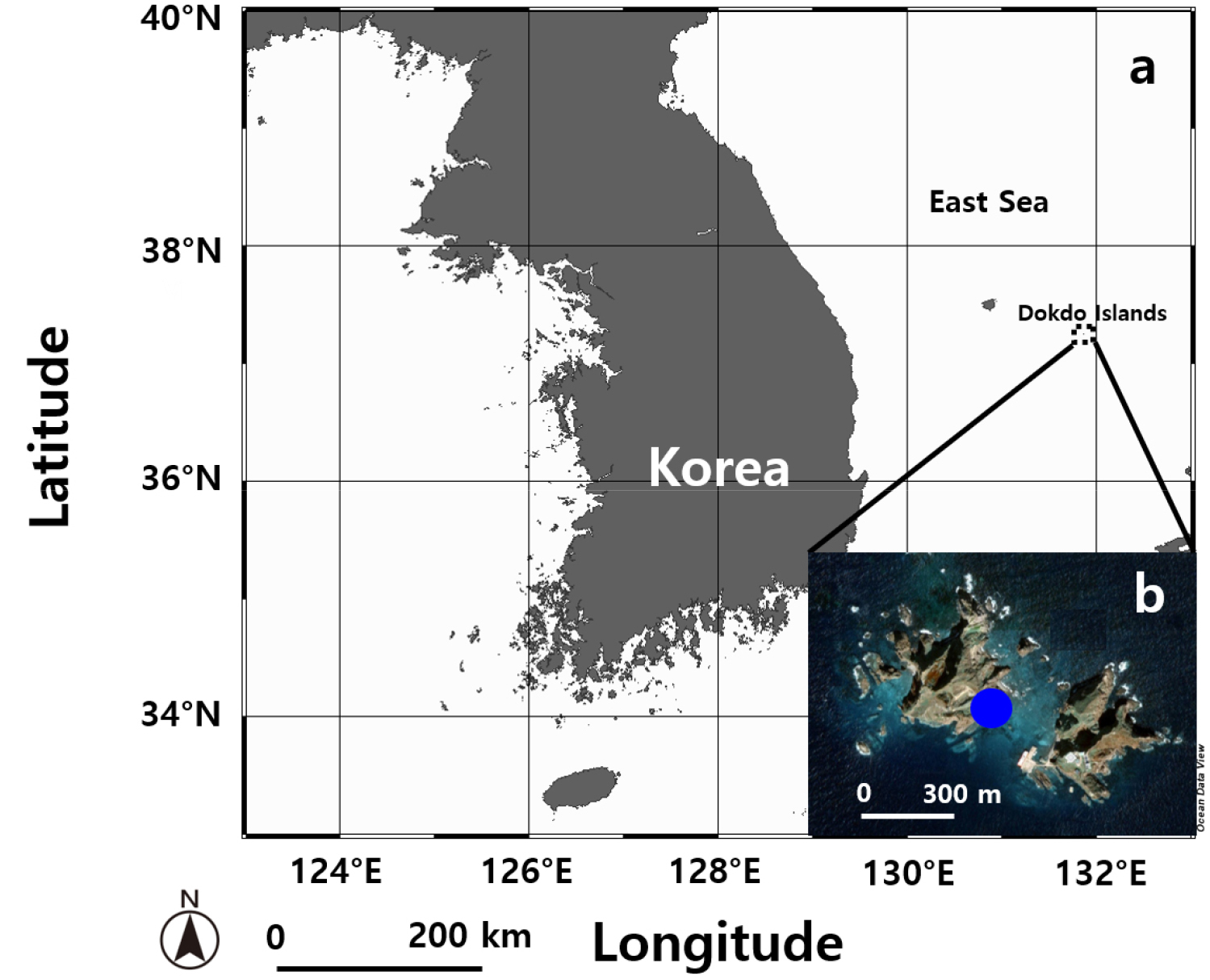
Fig. 1.
Map of the sampling site in the coastal area of the Dokdo Islands, Korea. (a) Overview map showing the location of the Dokdo Islands in the East Sea, southeast of Ulleungdo in Gyeongsangbuk-do. (b) Detailed image of the sampling site in the coastal region of Dokdo Islands, obtained from Google Earth.
Table 1.
Strain, location of collection (LC), water temperature (T, °C), salinity (S, psu), and GenBank accession number for marker genes of Tetraselmis marina MP0060 isolated from the Dokdo Islands in the East Sea of Korea
Morphological identification
The morphology of living cells grown photosynthetically at the exponential growth phase, approximately 2 × 105 cells mL-1, was examined using an inverted microscope (CKX53; Olympus, Tokyo, Japan). Cell length and width were measured for 30 randomly selected cells (n = 30) from the flagellated and non-motile stages using a digital camera (Zeiss AxioCam MRc5; Carl Zeiss, Göttingen, Germany). Results are presented as mean ± standard error to ensure statistical robustness.
For field emission scanning electron microscopy (FE-SEM), 10 mL aliquots of cultures at the exponential growth phase, with a density of approximately 2 × 105 cells mL-1 were fixed in a commercially available 4% (w/v) osmium tetroxide (OsO4; Electron Microscopy Sciences, Hatfield, PA, USA) solution, which was diluted to a final concentration of 1% (w/v) by mixing with the culture medium. The fixation was carried out for 10 min without using any buffer system. The fixed cells were collected on polycarbonate membrane filters with a pore size of 3 µm and a diameter of 25 mm (Whatman Nuclepore Track-Etched Membranes; Whatman, Kent, UK) and washed thrice with distilled water. The membranes with the attached cells were dehydrated in a graded ethanol series (10%, 30%, 50%, 70%, 90%, and 100%), followed by two changes in 100% ethanol (100% absolute alcohol; Merck, Darmstadt, Germany). Next, they were immediately dried using an automated critical point dryer (EM CPD300; Leica, Wetzlar, Germany) with CO2 for critical point drying. The dried filters were mounted on an aluminum stub (Electron Microscopy Sciences) using copper conductive double-sided tape (Ted Pella, Redding, CA, USA) and coated with gold using an ion sputter (MC1000; Hitachi, Tokyo, Japan). The cells and surface morphologies were observed using a high-resolution Zeiss Sigma 500 VP FE-SEM (Carl Zeiss).
Cells in the exponential growth phase, with a density of approximately 2 × 105 cells mL-1, were transferred to a 10 mL tube and fixed in 2.5% (v/v) glutaraldehyde (Electron Microscopy Sciences) (final concentration) for 1.5 h for transmission electron microscopy (TEM). The tube contents were placed in a 10 mL centrifuge tube and then centrifuged at 1610 × g for 10 min (VS-5500; Vision, Bucheon, Republic of Korea). The pellet was transferred to a 1.5 mL tube and rinsed several times with 0.2 M sodium cacodylate buffer at pH 7.4 (Electron Microscopy Sciences). The cells were post-fixed with 1% (w/v) OsO4 prepared in deionized water for 90 min. The pellet was embedded in agar before dehydration in a graded ethanol series (50%, 60%, 70%, 80%, 90%, and 100%), followed by two changes in 100% ethanol. The material was then embedded in Spurr’s resin (Electron Microscopy Sciences). Sections were prepared using an EM UC7 ultramicrotome (Leica) and stained with 3% (w/v) aqueous uranyl acetate (Electron Microscopy Sciences), followed by 0.5% (w/v) lead citrate (Electron Microscopy Sciences) (Reynolds 1963). The sections were visualized using TEM (Sigma 500/VP TEM; Carl Zeiss).
Molecular and phylogenetic analysis
For molecular analysis, genomic DNA (gDNA) was extracted from cells in the exponential growth phase, with a density of approximately 2 × 105 cells mL-1, using an AccuPrep Genomic DNA Extraction Kit (Bioneer, Daejeon, Republic of Korea) in accordance with the manufacturer’s instructions. The primers used to amplify each marker gene are listed in Table 2. The reaction mixtures for PCR amplification comprised 5 µL of 10× F-Star Taq reaction buffer, 1 µL of 10 mM dNTP mix, 0.02 µM of primers, 0.25 µL of 5 U µL-1 BioFACT F-Star Taq DNA polymerase (BioFACT Co., Ltd., Daejeon, Republic of Korea), 38.75 µL of UltraPure DNAse/RNAse-free distilled water (Invitrogen, Carlsbad, CA, USA), and 3 µL of the DNA template (ca. 10–30 ng DNA). PCR amplification was performed on an Eppendorf Mastercycler PCR machine (Eppendorf, Hamburg, Germany) under the following thermal cycling conditions: pre-denaturation at 94°C for 5 min, followed by 35 cycles of 94°C for 1 min, the selected annealing temperature (AT) for 1 min, 72°C for 1 min, and final extension at 72°C for 5 min. The ATs of the primer sets were determined through gradient PCR. We optimized the ATs as follows: 56°C (EukA-G18R), 56°C (570F-EukB), 57°C (ITSF2-ITSFR2), and 57°C (D1R-LSUB). PCR products were purified using the AccuPrep PCR Purification Kit (Bioneer) and subjected to Sanger sequencing (Macrogen, Daejeon, Republic of Korea). Nucleotide sequences were identified using the National Center for Biotechnology Information (NCBI) Basic Local Alignment Search Tool. Alignments and phylogenetic and molecular evolutionary analyses of the obtained sequences were conducted utilizing Geneious Prime v.2022.2.2. (Biomatters Ltd., Auckland, New Zealand). The analyses incorporated various assemblages, drawing on data from other species available in the NCBI GenBank database. Bayesian analyses were run using MrBayes v.3.2.7 (Huelsenbeck and Ronquist 2001; Ronquist and Huelsenbeck 2003) with the default GTR + G + I model to determine the best available model for the data of each region. Four independent Markov chain Monte Carlo runs were performed as described by Kang et al. (2010) for all sequence regions. Moreover, maximum-likelihood analyses were conducted using RAxML v.8.2.10 (Stamatakis 2014). Two hundred independent free inferences were allowed, and the –# option was used to identify the best tree. Bootstrap values were calculated with 1000 replicates using the same substitution model.
Table 2.
Primers used in this study to amplify the small-subunit (SSU) rDNA, ITS1-5.8S-ITS2 (ITS), and the 28S rRNA gene region of the large-subunit (LSU) rDNA in Tetraselmis marina MP0060
| Primer name | Primer Region | Sequence (5′–3′) |
Annealing Temperature (°C) |
Target Base Pair Length (bp) | Reference |
| EukA | Forward, SSU | AACCTGGTTGATCCTGCCAG | 56 | ~1,740 | Medlin et al. 1988 |
| G18R | Reverse, SSU | GCATCACAGACCTGTTATTG | 56 | ~1,740 | Litaker et al. 2003 |
| 570F | Forward, SSU | GTAATTCCAGCTCCAATAGC | 56 | ~1,740 | Weekers et al. 1994 |
| EukB | Reverse, SSU | TGATCCTTCTGCAGGTTCACCTAC | 56 | ~1,740 | Medlin et al. 1988 |
| ITSF2 | Forward, ITS | TACGTCCCTGCCCTTTGTAC | 57 | ~578 | Litaker et al. 2003 |
| ITSFR2 | Reverse, ITS | TCCCTGTTCATTCGCCATTAC | 57 | ~578 | Litaker et al. 2003 |
| D1R | Forward, LSU | ACCCGCTGAATTTAAGCATA | 57 | ~1,238 | Scholin et al. 1994 |
| LSUB | Reverse, LSU | ACGAACGATTTGCACGTCAG | 57 | ~1,238 | Litaker et al. 2003 |
Determination of optimal culture conditions
Routine serial sub-culturing was performed in fresh F/2 medium to maintain a pure culture of MP0060. For the experiments, a portion of this strain was inoculated into F/2 medium at an initial cell density of 5 × 103 cells mL-1 and incubated at 22°C under cool white fluorescent light (~60 µmol photons m-2 s-1, PAR) with a 14:10 h light/dark cycle for 21 days. The 21-day incubation period was conducted to grow the culture to the exponential growth phase before initiating the optimal culture test. After incubation, the culture reached the exponential growth phase with a density of approximately 2 × 105 cells mL-1. Subsequently, the culture was used to conduct an optimal culture test at a laboratory scale.
Optimal temperature and illumination analyses were conducted simultaneously using a PhotoBiobox (Heo et al. 2015). The culture was adjusted to a concentration of 1 × 105 cells mL-1 using F/2 medium and 200 µL of the adjusted culture was dispensed into each well of 96-well black/clear bottom plates (Thermo Fisher Scientific, Waltham, MA, USA). These well plates were designed with black sidewalls to minimize cross-contamination and clear bottoms to enhance light penetration. After dispensing the culture, the plates were sealed with an optically clear well plate sealing film (Thermo Fisher Scientific) to prevent evaporation and contamination.
After incubation for 72 h in PhotoBiobox controlled at 5–40°C and 0–350 µmol m-2 s-1, optical density was measured at 600 nm using a Synergy II microplate reader (Biotek, Winooski, VT, USA). The OD at 600 nm was chosen to minimize interference from pigments and was used as an indirect measure of biomass. The initial cell density at the start of the experiment was adjusted to 1 × 105 cells mL-1 to ensure consistent growth across all conditions. The experiment was conducted in triplicate to enhance reproducibility. No significant deviations in OD values were observed across replicates, indicating stable growth during the experiment. The specific growth rate (µ) was calculated using the formula µ = (ln A2 - ln A1)/(T2 - T1), where A1 and A2 represent the O.D. values at T = 0 and T = 72 h, respectively. The calculated growth rates were visualized as heat maps using Microsoft Excel 2019 (Microsoft, Redmond, WA, USA).
Analyses for fatty acid composition of lipids
Lipid extraction was performed using a modified version of the Bligh–Dyer method, as described by Breuer et al. (2013), after 21 days of culture in F/2 medium under photoautotrophic conditions at 21°C and 88 µmol photons m-2 s-1 (PAR) with a 14:10 h light/dark cycle, which were determined as the optimal conditions for growth. At the time of lipid extraction, the culture was in the exponential growth phase, with a density of approximately 2 × 105 cells mL-1. Composition was analyzed using a 7890A gas chromatograph equipped with a 5975C mass selective detector (Agilent Technologies, Santa Clara, CA, USA) as described in our previous study (Kang et al. 2019). Compound identification was performed by matching the mass spectra with those in the Wiley/NBS registry of the mass spectral data. A search with a match value greater than 90% was considered valid.
3. Results and Discussion
Morphological characteristics
Under light microscopy, MP0060 exhibited three distinct stages, namely, flagellate, non-motile, and cyst, each characterized by unique morphological features (Fig. 2). In the flagellated stage, cells exhibited an elliptical to oval shape when viewed from the ventral side (Fig. 2a–c). The ranges (mean ± standard error, n = 30) of the cell lengths and widths in this stage were 11.0–16.5 µm (13.5 ± 0.3) and 5.4–8.8 µm (7.5 ± 0.1), respectively. The cells in this stage were green, containing a single large, cup-shaped chloroplast that was typically located at the cell's periphery (Fig. 2a and b) and occasionally toward the posterior end. A centrally located pyrenoid was observed within the chloroplast, and a conspicuous orange-red eyespot (stigma) was situated in the anterior part of the cell (Fig. 2b). The cells were located at the anterior end of a depression (Fig. 2b), from which four uniform flagella extended through an anterior cleft, each flagellum bearing fine hairs (Fig. 2c). Additionally, in the non-motile stage (Fig. 2d and e), colony-forming cells developed a stalk composed of transversely septate compartments (SC1–SC4). Each compartment was empty, with terminal cells at the ends (Fig. 2e). The length and width of these compartments were (mean ± standard error, n = 30) 8.5–15.6 µm (11.7 ± 0.3) and 4.3–6.0 µm (5.2 ± 0.1), respectively. Cells in the cyst stage appeared circular in ventral view, surrounded by distinct, thick cell walls.The diameter of the cysts was (mean ± standard error, n = 30) 11.1–18.3 μm (14.7 ± 0.5 μm) (Fig. 2f).
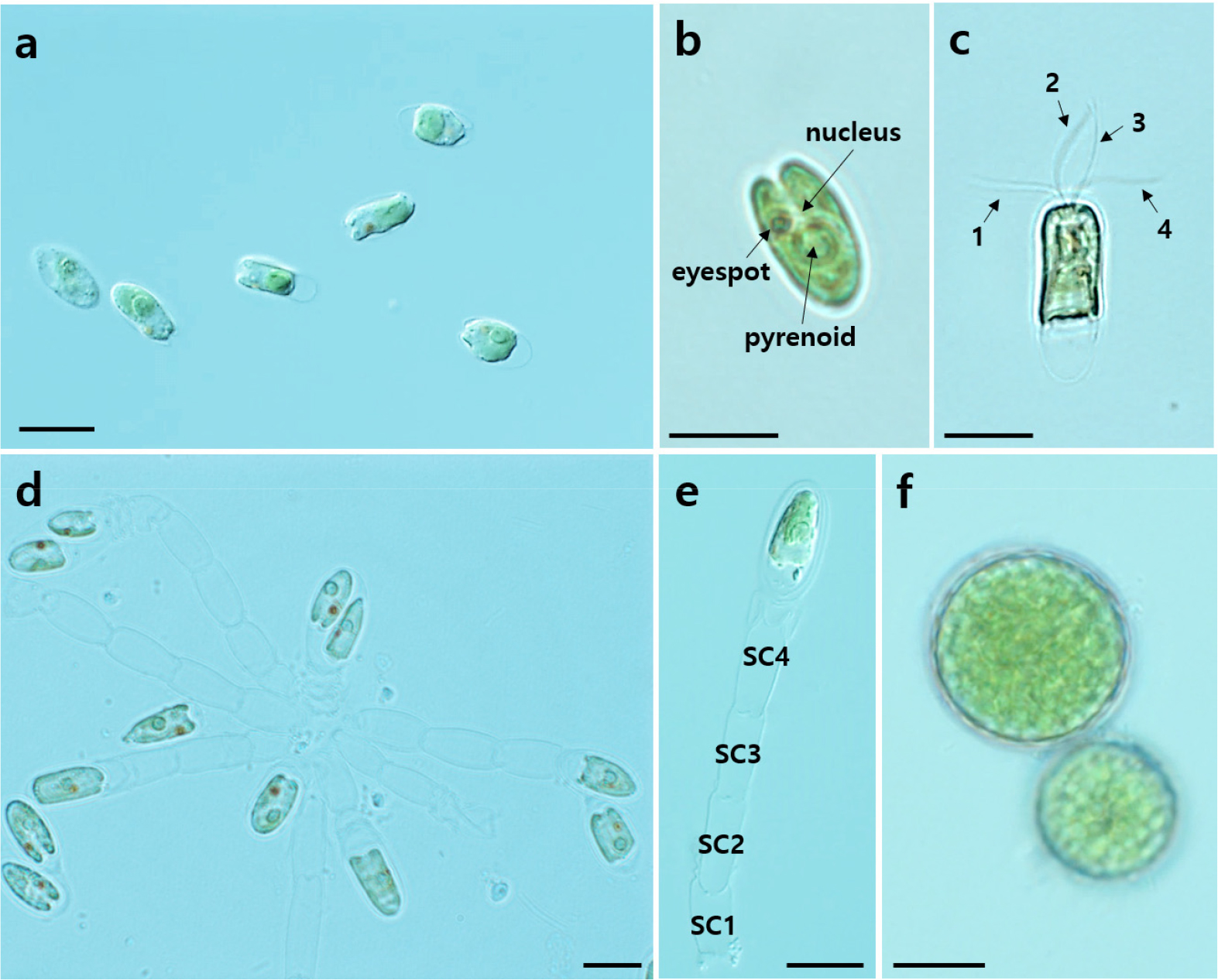
Fig. 2.
Light micrographs of Tetraselmis marina MP0060 illustrating three distinct stages: flagellate (a–c), non-motile (d–e), and cyst (f). (a) Flagellate cells viewed from the ventral side, showing an elliptical to oval shape. (b) Flagellate cell with a centrally located pyrenoid and a prominent orange-red eyespot in the anterior region. The nucleus is positioned between the anterior and central parts. (c) Flagellate cell with four flagella (labeled 1–4) extending through an anterior cleft. (d) Non-motile stage showing cells forming colonies with a stalk structure. (e) Micrographs showing the stalk with transversely septate compartments (labeled SC1–SC4) and terminal cells at the ends. (f) Cyst formation stage, with cells appearing circular and surrounded by a distinct, thick cell wall. Scale bars: a–f = 10 µm.
Scanning electron micrographs of MP0060 revealed three distinct stages, namely, flagellate, non-motile, and cyst, each exhibiting unique morphological characteristics, as shown in Fig. 3. In the flagellate stage, cells displayed an elliptical shape with a smooth surface texture when viewed from the ventral side (Fig. 3a). Additionally, Fig. 3b illustrates four flagella (labeled 1–4) emerging from an apical depression at the base of the cell.
In the non-motile stage (Fig. 3c–e), the colonial form of MP0060 exhibited elongated stalks composed of multiple transversely septate (Fig. 3c and e), slightly barrel-shaped compartments (Fig. 3c and e), each of which was not enclosed within a common theca (Fig. 3d and e). The stalk structure typically contained two to ten compartments, culminating in a terminal cell with distinct surface ridges (Fig. 3d). In the cyst formation stage, cells appeared nearly spherical, featuring an ornamented cyst wall with a dense array of spine-like structures on the surface (Fig. 3f).
Transmission electron micrographs of MP0060 revealed three life stages, namely, flagellate, non-motile, and cyst, each displaying key ultrastructural features (Fig. 4). Thin sections prepared for TEM highlighted primary cellular components, such as the chloroplast (ch), eyespot (e), Golgi apparatus (g), mitochondria (m), nucleus (n), pyrenoid (p), and starch (s) (Fig. 4a–d).
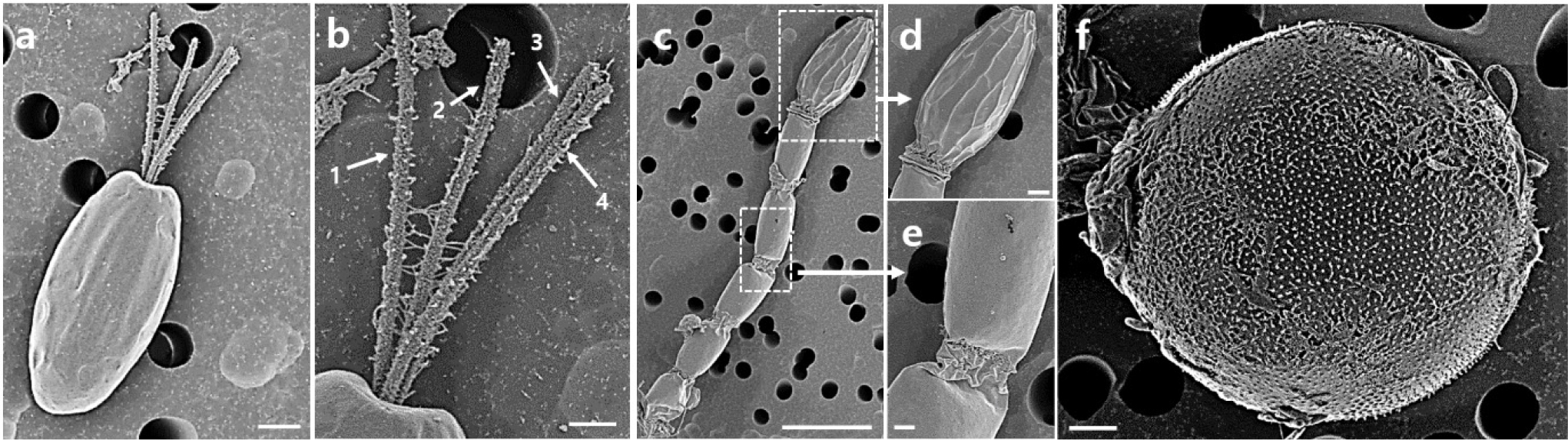
Fig. 3.
Scanning electron micrographs of Tetraselmis marina MP0060 showing three distinct stages: flagellate (a–b), non-motile (c–e), and cyst (f). (a) Flagellate cell displaying an elliptical shape with a smooth surface texture, viewed from the ventral side. (b) Detailed view showing four flagella (labeled 1–4) emerging from an apical depression at the cell base. (c) Non-motile stage in colonial form, exhibiting elongated stalks composed of multiple transversely septate, slightly barrel-shaped compartments. (d) Enlarged view of the terminal cell in the stalk, showing distinct surface ridges. (e) Close-up of the stalk compartments. (f) Cyst stage, with cells appearing nearly spherical and featuring an ornamented cyst wall covered with spine-like structures. Scale bars: c = 10 µm, a, e & f = 2 µm, b & d = 1 µm.
In the flagellate stage, chloroplasts were predominantly located along the cell periphery, with starch grains near the pyrenoid matrix (Fig. 4a and c). Two distinct starch deposits were observed: solitary grains within the chloroplast stroma and clusters surrounding the pyrenoid matrix. The pyrenoid matrix (p) was enclosed by starch grains (s), positioned near the nucleus (n) as shown in Fig. 4a. The flagellar apparatus included four basal bodies (b1–b4) in a zig-zag arrangement (Fig. 4b), with rhizoplasts (r) adjacent to them. Additionally, the Golgi apparatus (g) was situated near the basal bodies on both sides of the flagellar base (Fig. 4b). The eyespot (e) appeared at the cell periphery, with lipid granule layers separated by a thylakoid, parallel to the chloroplast membrane (Fig. 4c and d).
In the non-motile stage, individual and paired cells with distinctive protoplast structures were observed. Fig. 4e shows a single-cell structure, Fig. 4f a paired structure, and Fig. 4g two separate paired structures. Some cells were surrounded by multiple closely arranged thecal walls, forming a compact structure around the protoplast (Fig. 4h). The cells in this stage retained ultrastructural features identical to the motile cells, with preserved organelles and internal structures.
The cyst stage featured a thick wall and numerous surface spines (Fig. 4i), with ultrastructural elements such as chloroplasts (ch), pyrenoid (p), nucleus (n), starch grains (s), and vacuole (v) also preserved.
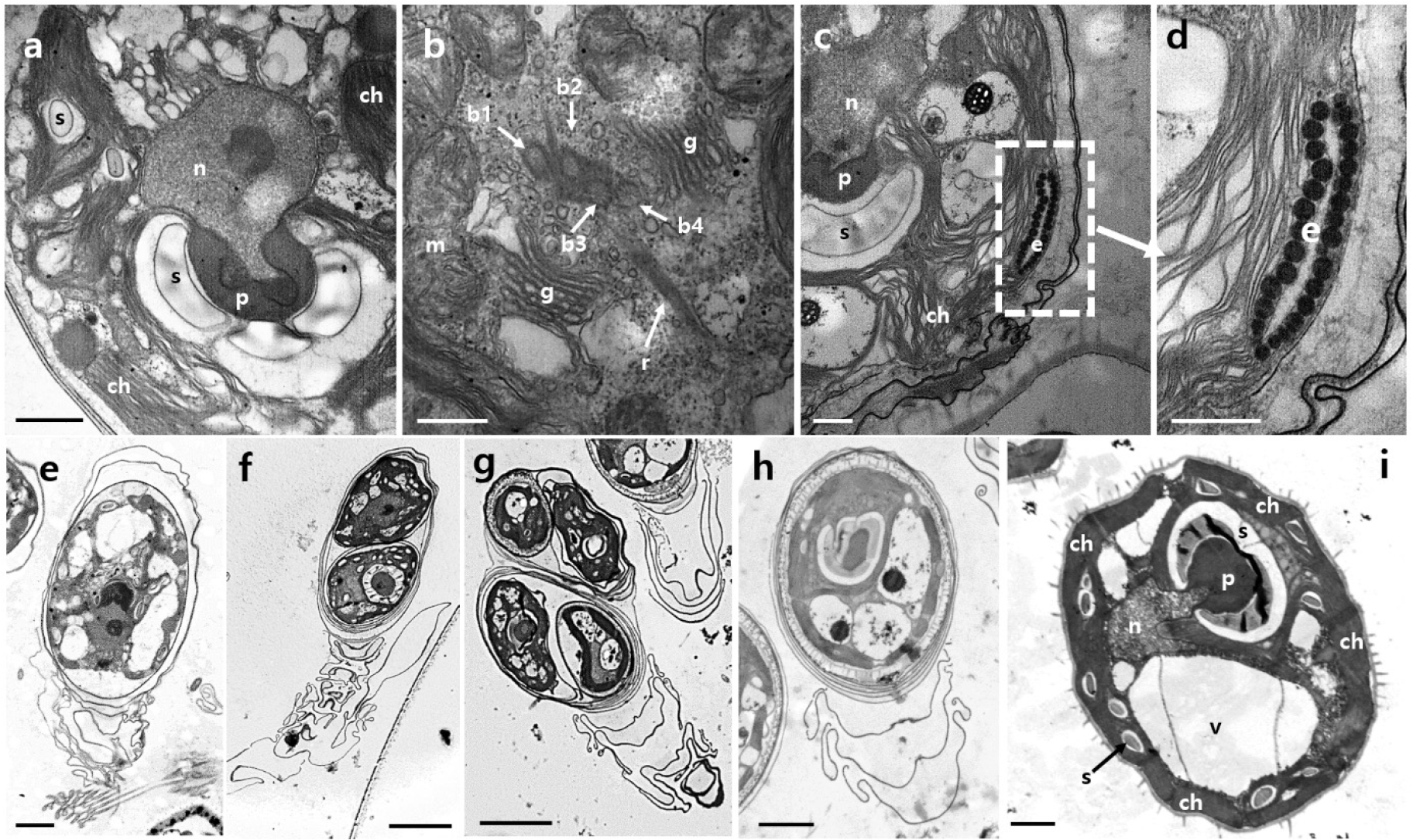
Fig. 4.
Transmission electron micrographs (TEM) of Tetraselmis marina MP0060 illustrating ultrastructural features across three life cycle stages: flagellate (a–d), non-motile (e–h), and cyst (i). (a) Flagellate cell showing main features of the cell, including the chloroplast (ch), nucleus (n), pyrenoid (p), and starch grains (s). (b) Flagellar apparatus with four basal bodies (b1–b4) in a zig-zag arrangement, accompanied by rhizoplasts (r), mitochondria (m), and the Golgi apparatus (g) adjacent to the flagellar base. (c) Flagellate cell displaying the chloroplast (ch) with starch deposits near the pyrenoid (p) and a peripheral eyespot (e). (d) Enlarged view of the eyespot (e) showing lipid granule layers separated by a thylakoid, parallel to the chloroplast membrane. (e) Non-motile stage showing a single-cell structure. (f) Non-motile stage showing a paired cell structure. (g) Non-motile stage showing two paired cell structures. (h) Non-motile cell with multiple thecal layers surrounding the protoplast. (i) Cyst stage cell featuring a thick wall with numerous surface spines, containing ultrastructural elements such as the chloroplast (ch), pyrenoid (p), nucleus (n), starch grains (s), and vacuole (v). Scale bars: f & g = 5 µm, e & h = 2 µm, a & i = 1 µm, b–d = 0.5 µm.
The morphological characteristics of MP0060 closely aligned with the established description of the genus Tetraselmis. The compressed cell shape, with four flagella of equal length emerging from a slit at the apical base, is characteristic of this genus. Additionally, MP0060 possesses a single large, cup-shaped chloroplast containing one central pyrenoid, a prominent eyespot, and a nucleus positioned parallel to the pyrenoid and flagellar base. These features are consistent with the typical morphological traits described for Tetraselmis, further confirming the classification of MP0060 within this genus (Norris et al. 1980; Hori et al. 1983; Hyung et al. 2021). Furthermore, MP0060 exhibited morphological and ultrastructural characteristics consistent with those reported for T. marina species across various regions, including Europe (Parke and Manton 1965), North America (Hori and Chihara 1974), and Japan (Hori et al. 1983), as summarized in Table 3. While this study focused on observing the three distinct morphological stages (flagellate, non-motile, and cyst), the environmental triggers inducing these lifecycle transitions were not specifically investigated. Nevertheless, the observed stages align well with the lifecycle patterns described for T. marina (Norris et al. 1980), where motile cells act as dispersal units before transitioning to non-motile stages under suitable substrate conditions. The non-motile stage, characterized by the formation of a cellular stalk with transversely septate compartments, likely facilitates colony stability and adaptation to ecological niches. Finally, the cyst stage, with its thick ornamented walls, highlights a mechanism for environmental resistance. A distinctive feature differentiating this isolate from other species within the genus is the formation of a cellular stalk. The cellular stalk, as reported in previous studies, represents an adaptive feature that distinguishes T. marina from other Tetraselmis species, playing a crucial role during the non-motile stage in colony establishment and persistence (Norris et al. 1980; Table 3). These shared characteristics across multiple studies provide conclusive evidence for the identification of our isolate as T. marina. Further studies investigating the specific environmental parameters, such as nutrient availability, light intensity, and substrate type, that regulate lifecycle transitions in MP0060 could offer deeper insights into the adaptive strategies of this species. Such investigations would complement the morphological observations presented here and clarify the ecological role of stalk formation and cyst development.
Table 3.
Comparison of morphological and ultrastructural characteristics among Tetraselmis marina strains and Tetraselmis jejuensis
| Morphological and Ultrastructural Traits | Tetraselmis marina MP0060 (Korea) |
Reported Strains (Global) | Tetraselmis jejuensis |
| Strain locality | Reported from Korea | Widely reported from Europe, North America, Japan, and other temperate regions | Reported from Korea |
| Cell shape | Elliptical | Elliptical | Elliptical |
|
Cell length (µm; flagellated stage) | 11.0–16.5 (mean: 13.5) | 16–20 | 13.0-20.8 (mean: 16.7) |
|
Cell width (µm; flagellated stage) | 5.4–8.8 (mean: 7.5) | 7–8 | 6.5-16.3 (mean: 11.3) |
|
Cell width (µm; non-motile stage, colonial) | 4.3–6.0 (mean: 5.2) | 6–8 | ND |
|
Cell size (µm; cyst formation stages) |
11.1–18.3 (mean: 14.7) in diameter | ND | ND |
| Flagella | Four flagella | Four flagella | Four flagella |
| Nucleus | Large, located near the center of the cell | Large, located near the center of the cell | located in the upper part of the cell |
| Chloroplast | Cup-shaped | Cup-shaped | Cup-shaped |
| Pyrenoid | Present, large, located below the nucleus | Present, large, located below the nucleus | Present, large, located below the nucleus |
| Starch grains | Surround the pyrenoid, concave on the side adjacent to the pyrenoid | Surround the pyrenoid, concave on the side adjacent to the pyrenoid | Surround the pyrenoid, concave on the side adjacent to the pyrenoid |
| Eyespot | Present | Present | Present |
| Colonial structure | Forms colonies with a septate stalk, barrel-shaped | Forms colonies with a septate stalk, barrel-shaped | ND |
| Reference | This study | Hori et al. 1983 | Hyung et al. 2021 |
Molecular identification and sequence analysis
In this study, the combined length of the SSU, ITS1- 5.8S-ITS2, and LSU rDNA sequences in the newly isolated strain was found to be 3,556 nucleotides, with GenBank accession numbers KY655019–KY655021, as detailed in Table 1. The alignment results revealed that the SSU rDNA sequence of MP0060 closely matched those of strains CCMP898 (Whidbey Island, USA), IMA043 (Chioggia, Italy), and KNUA020 (Ría de Vigo, Spain), each showing a single base pair difference (0.1%). Strain AC16-MESO (Antofagasta coast, Chile) exhibited three base pair differences (0.2%), followed by strain BEA0158B (Gran Canaria, Spain) with 15 base pair differences (0.9%). Strain CTM 20015 (Kelibia, Tunisia) showed 17 base pair differences (1.0%), whereas the most significant divergence was observed in strain NZmm1W1 (Naozhou Island, China), with 23 base pair differences (1.3%) (Table 4). Additionally, for the ITS region, strains IOAC331S and AC16-MESO (Antofagasta coast, Chile) showed no base pair differences. Strain CCMP898 (Whidbey Island, USA) exhibited one base pair difference (0.2%), whereas strain SAG 202.8 showed nine base pair differences (1.6%), as detailed in Table 4.
Table 4.
Comparison of small-subunit rDNA (SSU) and ITS1-5.8S-ITS2 (ITS) sequences of Tetraselmis marina MP0060 and other strains
| Strain name | Collection location | GenBank accession number | Tetraselmis marina MP0060* | Marker Gene |
| CCMP898 | Whidbey Island, USA | HE610131 | 1 (0.1) | SSU |
| IMA043 | Chioggia, Italy | OM319688 | 1 (0.1) | SSU |
| KNUA020 | Ría de Vigo, Spain | KY045847 | 1 (0.1) | SSU |
| AC16-MESO | Antofagasta coast, Chile | MN304931 | 3 (0.2) | SSU |
| BEA0158B | Gran Canaria, Spain | OQ220342 | 15 (0.9) | SSU |
| CTM 20015 | Kelibia, Tunisia | KT023599 | 17 (1.0) | SSU |
| NZmm1W1 | Naozhou Island, China | KY054995 | 23 (1.3) | SSU |
| IOAC331S | ND | KC800942 | 0 (0) | ITS |
| AC16-MESO | Antofagasta coast, Chile | KX752433 | 0 (0) | ITS |
| CCMP898 | Whidbey Island, USA | HE610131 | 1 (0.2) | ITS |
| SAG 202.8 | ND | MK460479 | 9 (1.6) | ITS |
Based on the phylogenetic analysis of SSU rDNA sequences, MP0060 grouped within a major T. marina clade, clustering closely with KNUA020, IMA043, AC16-MESO, CCMP898, CTM 20015, BEA0158B, and NZmm1W1 (Fig. 5). Similarly, in the phylogenetic tree derived from ITS1-5.8S-ITS2 rDNA sequences, MP0060 formed a clade with SAG 202.8, CCMP898, IOAC331S, and AC16-MESO, further supporting its placement within the T. marina lineage (Fig. 6). On the basis of the morphological and genetic results, this green alga was identified as T. marina. Accordingly, this microalga, designated as MP0060, is preserved as a live culture under the accession number MABIK LP00000080 at the National Marine Biodiversity Institute of Korea (MABIK). The SEM samples (MABIK FL00031099) and live cultures are also accessible at MABIK, ensuring their availability for future research.
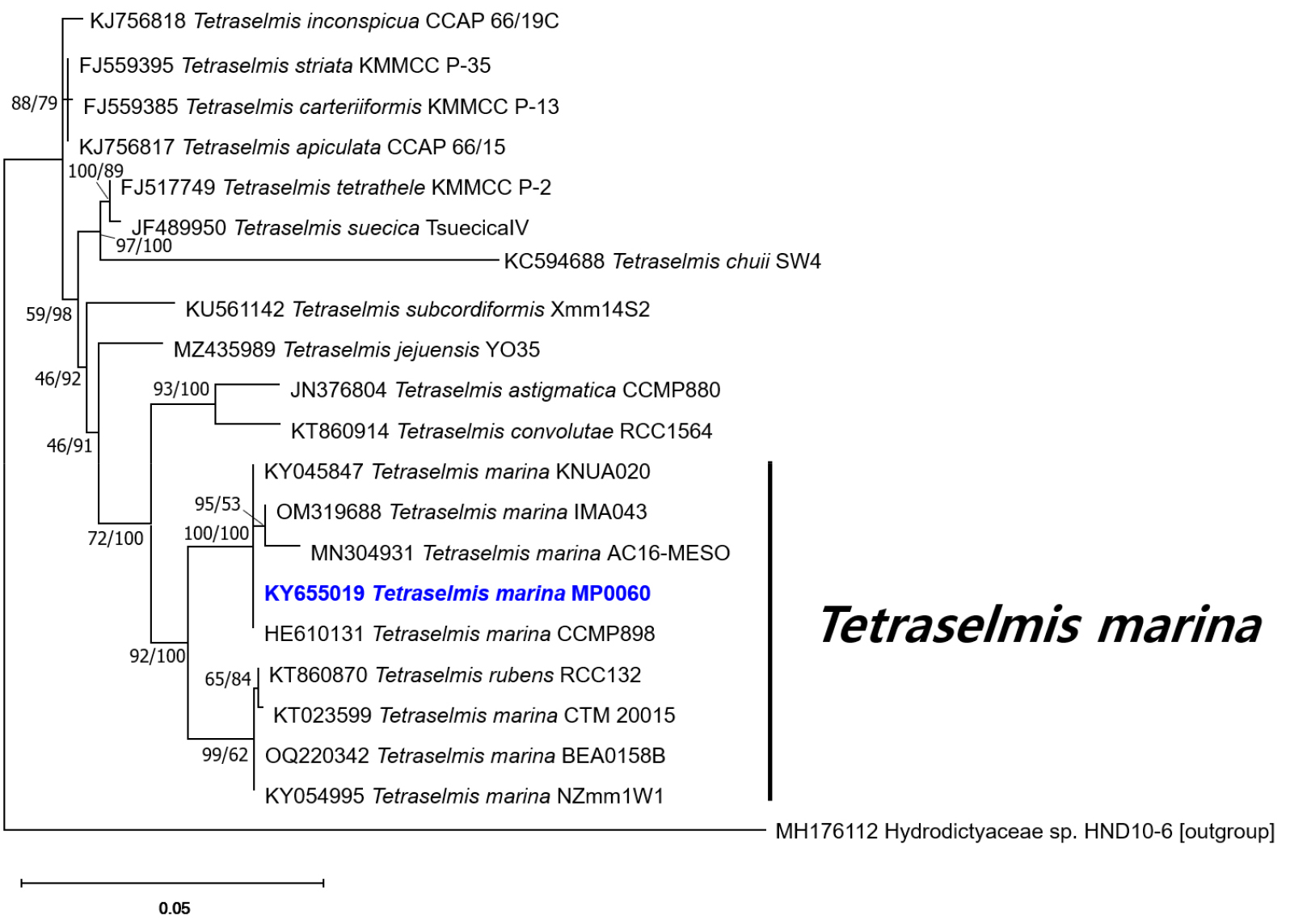
Fig. 5.
Maximum likelihood and Bayesian inference phylogenetic tree based on 18S rDNA sequences. The values on each node indicate maximum likelihood bootstrap percentages and Bayesian posterior probabilities (%). Tetraselmis marina MP0060 is highlighted in bold blue. The outgroup used is Hydrodictyaceae sp. HND10-6
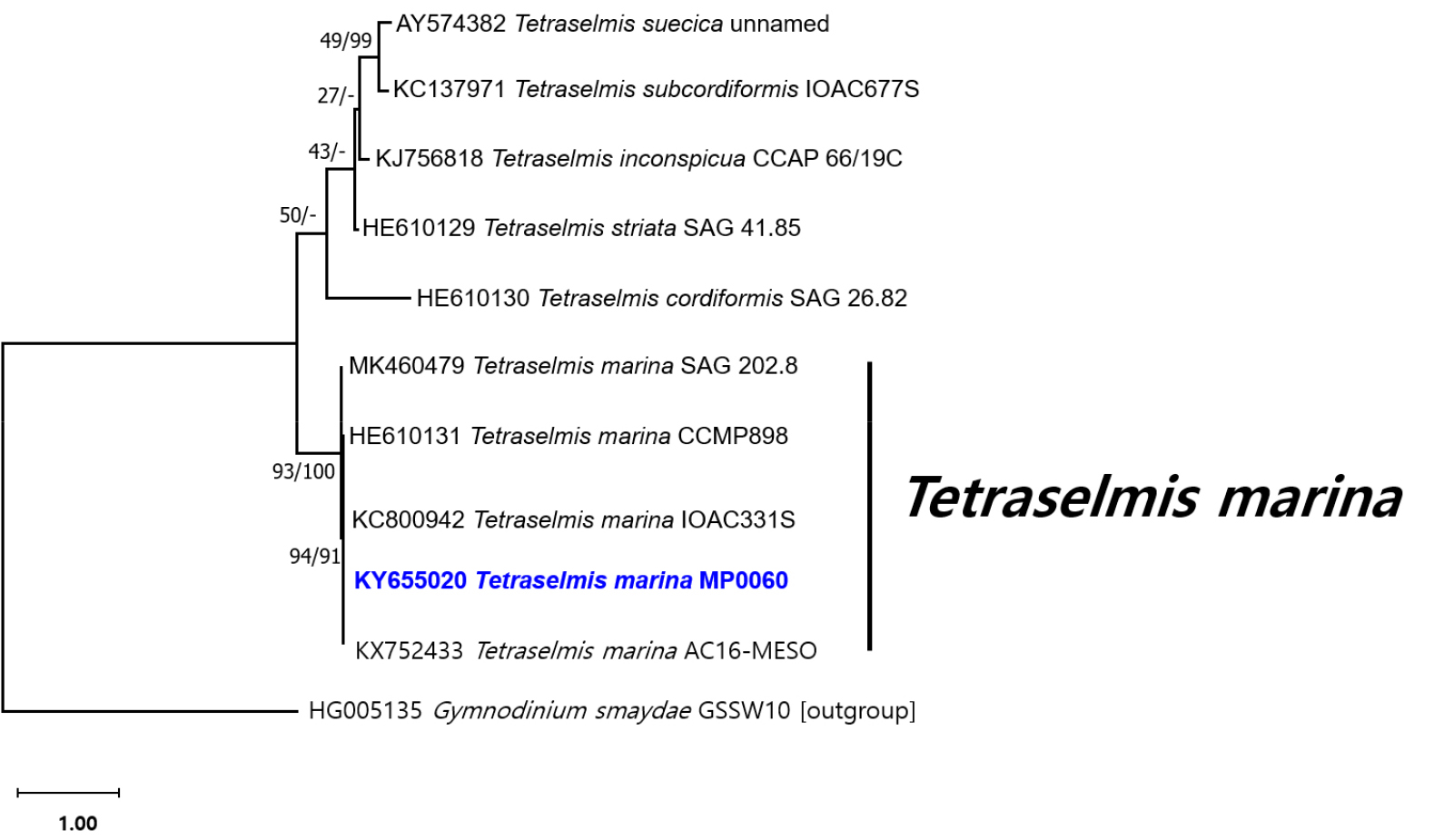
Fig. 6.
Maximum likelihood and Bayesian inference phylogenetic tree based on ITS1-5.8S-ITS2 rDNA sequences. The values on each node indicate maximum-likelihood bootstrap percentages and Bayesian posterior probabilities (%), respectively. Tetraselmis marina strain MP0060 is highlighted in bold blue. The outgroup used is Gymnodinium smaydae GSSW10
Verification of the optimal cultivation conditions of the isolated strain
Growth responses of MP0060 to a range of temperatures and light intensities were evaluated under laboratory- scale conditions to identify optimal cultivation parameters. As shown in Fig. 7, MP0060 exhibited growth across temperatures from 8 to 30°C, with the highest rates observed between 15 and 27°C under photon flux densities of 88–245 µmol m-2 s-1 (white LED). Optimal growth conditions were determined to be 21°C and 88 µmol m-2 s-1. With its adaptability to varying light intensities and temperatures, MP0060 demonstrated strong potential for growth across diverse environmental conditions. This flexibility in light and temperature likely supports its ability to thrive in a range of ecosystems, possibly explaining its broad geographic distribution, as indicated in Table 4. Such adaptability underscores the strain’s promise for numerous biotechnological applications, making it especially suited to industrial-scale production. Optimizing cultivation conditions and enhancing genetic traits could increase the viability of MP0060 as a valuable industrial resource.
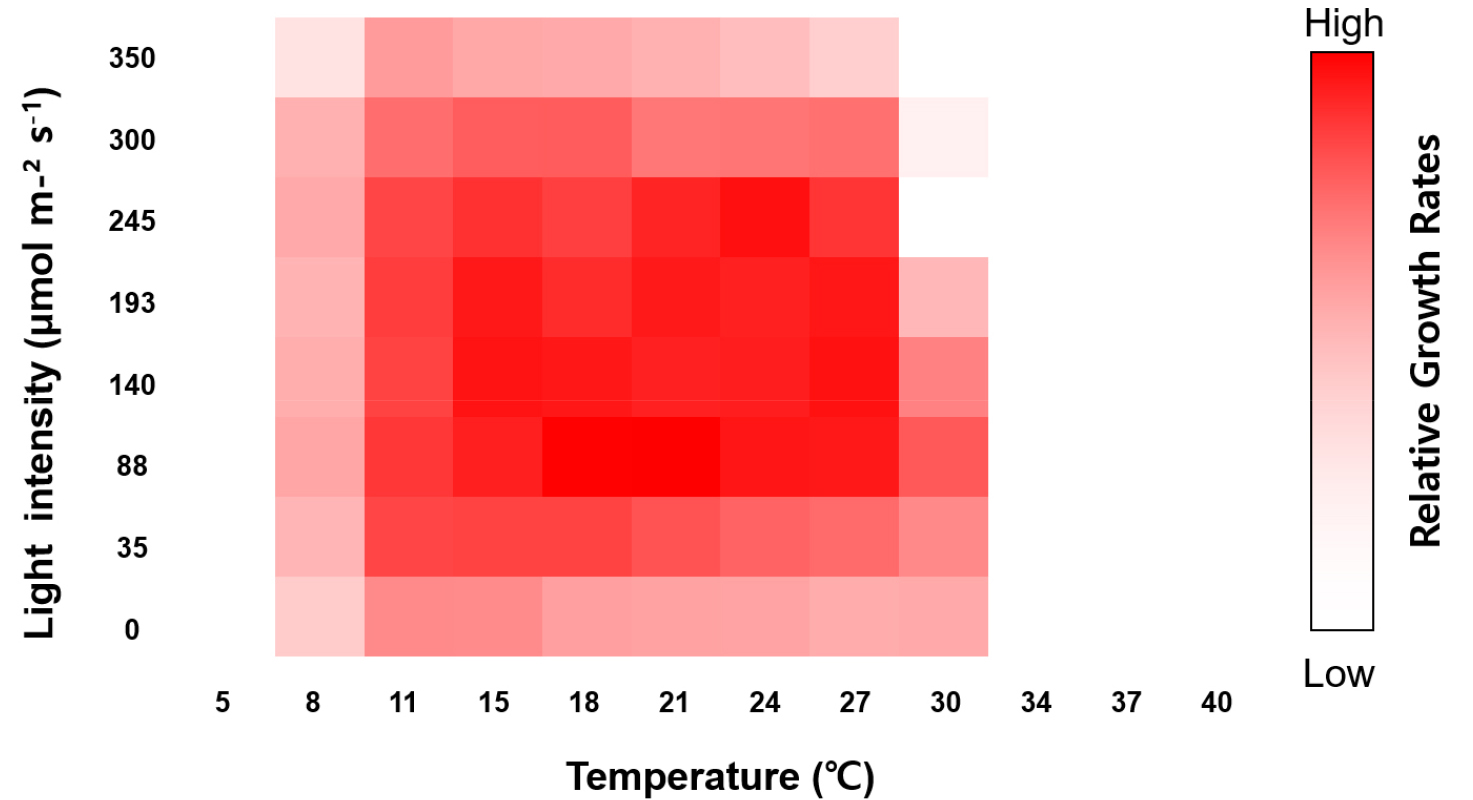
Fig. 7.
Heat map representing the relative growth rates of Tetraselmis marina MP0060 under varying light intensities (µmol m-2 s-1) and temperatures (°C), calculated from optical density (OD) measurements at 600 nm over a 72-hour incubation period. Higher growth rates are represented by more intense colors
Fatty acid composition of lipids
The fatty acid methyl ester (FAME) profiles of MP0060 revealed significant concentrations of palmitic acid (C16:0, 41.84%), oleic acid (C18:1 n-9, 10%), α-linolenic acid (C18:3 n-3, 17.49%), and eicosapentaenoic acid (C20:5 n-3, 14.04%) (Table 5). Additionally, stearic acid (C18:0, 5.4%) and eicosenoic acid (C20:1 n-9, 5.4%) were detected, accounting for the remaining fraction of the total fatty acid content. Saturated fatty acids (SFAs) accounted for 47.24% of the total fatty acids, whereas monounsaturated and polyunsaturated fatty acids (PUFAs) accounted for 15.4% and 37.36%, respectively (Table 5).
Table 5.
Lipid profile of Tetraselmis marina MP0060
| Component | Content (%)* | Note |
| Palmitic acid (C16:0) | 41.84 ± 0.4 | SFA (major) |
| Stearic acid (C18:0) | 5.4 ± 0.3 | - |
| Oleic acid (C18:1 n-9) | 10 ± 0.4 | - |
| Linoleic acid (C18:2 n-6) | 5.83 ± 0.6 | - |
| α-linolenic acid (C18:3 n-3) | 17.49 ± 0.2 | ω-3 PUFA (major) |
| Eicosenoic acid (C20:1 n-9) | 5.4 ± 0.1 | - |
| Eicosapentaenoic acid (C20:5 n-3) | 14.04 ± 0.3 | ω-3 PUFA (major) |
| Total saturated fatty acids | 47.24 | |
| Total monounsaturated fatty acids | 15.4 | |
| Total polyunsaturated fatty acids | 37.36 |
The palmitic acid (C16:0) content of this strain accounts for 41.84% of its total fatty acids, making MP0060 a promising candidate for biodiesel production. This is because SFAs like palmitic acid improve the oxidative stability of biodiesel, enhancing its resistance to oxidation and degradation (Talebi et al. 2013; Díaz et al. 2014).
In comparison to other species within the same genus, T. marina MP0060, cultured under optimal conditions of 21°C and 88 µmol photons m-2 s-1, shows a significantly higher palmitic acid concentration than T. suecica P03A (36.8%) and T. chuii SW4 (24.98%) (Viso and Marty 1993; Andrew et al. 2022). Even when compared with other T. marina strains, such as P02D (28.8%) and AC16-MESO (22.5%) (Viso and Marty 1993; Mata et al. 2023), MP0060 remains to have considerably higher palmitic acid content. The distinct lipid profile of MP0060, with a high palmitic acid concentration and elevated SFA content, underscores its strong potential as a biodiesel feedstock. This composition makes it particularly suitable for biofuel applications, where enhanced oxidative stability is essential for industrial use.
Essential PUFAs, such as α-linolenic acid (ALA) and eicosapentaenoic acid (EPA), play crucial roles in regulating inflammation, promoting cardiovascular health, and preventing cancer (Simopoulos 2002).
Compared with other strains within the genus, MP0060 shows higher levels of ALA (17.49%) and EPA (14.04%). For instance, compared with MP0060, P03A recorded lower ALA (11.5%) and EPA (4.2%) contents, whereas other T. marina strains such as P02D (ALA 16%, EPA 5.8%) and AC16-MESO (ALA 9.1%, EPA 6.1%) showed comparatively lower values (Viso and Marty 1993; Mata et al. 2023). Such a profile suggests that MP0060, grown under the specified optimal conditions, could serve as a potent source of essential PUFAs, offering substantial potential in nutraceutical applications.
In conclusion, the present study provides the first report of T. marina MP0060 from the Dokdo Islands in the East Sea of Korea, marking the first documentation of this species in Korea and representing an important contribution to the documented marine biodiversity of the region. Comprehensive morphological and molecular analyses of this strain established its place within the Tetraselmis genus and provided insights into its unique features. Additionally, we identified the optimal culture conditions and fatty acid profile of MP0060. Specifically, MP0060 had notably high palmitic acid and PUFA contents, underscoring its potential for industrial applications in biofuels and essential fatty acids. A clonal culture of MP0060 has been successfully established and preserved in a national culture collection, ensuring accessibility for future studies and applications. These findings not only enhance the taxonomic understanding of T. marina but also open opportunities for its utilization in sustainable industrial applications.
