

1. 서 론
2. 재료 및 방법
조사 해역
기후요인
현장 환경요인 측정
중형동물플랑크톤 채집과 분석
통계분석
3. 결 과
기후요인
독도 환경요인
계절별 중형동물플랑크톤 군집특성
계절별 중형동물플랑크톤 군집 변화와 환경요인 간의 관계 분석
정준상관분석
4. 고 찰
계절별 동물플랑크톤 군집특성
기후변동과 동물플랑크톤 특성
5. 결 론
1. 서 론
전 지구적인 기후변동은 서로 다른 수준의 다양한 요인을 통해 해양생태계에 영향을 끼친다(Beaugrand et al. 2009; IPCC 2019). 수온, 염분, 용존산소, 그리고 먹이생물을 포함한 환경요인 중에서 수온은 동물플랑크톤에 가장 직접적인 영향을 미치는 것으로 알려져 있다(Hays et al. 2005; Richardson 2008; Moon et al. 2020). 수온은 동물플랑크톤 성장과 생존에 직접적인 영향을 줄 뿐만 아니라, 수층 밀도와 성층구조, 그리고 피-포식 관계에 영향을 미쳐서 동물플랑크톤 분포(종 구성과 개체수) 변화를 야기한다(Mackas et al. 2012; Doan et al. 2019). 우리나라 해역의 평균 표층수온은 지난 57년(1968–2024년)간 약 1.58°C정도 상승하여 전 지구 평균 대비 약 2배 이상 높았고, 동해의 경우 평년 대비 2–6°C 정도 높게 나타나 전 세계에서 평년 대비 가장 높았다(국립수산과학원 2022; 2025). 이와 같은 기후 및 해양환경의 변동은 플랑크톤 군집의 조성과 개체수 변동을 통해 해양 생태계 전반의 구조와 기능에 영향을 미칠 수 있다.
동해는 한반도 주변 해역 중 표층 수온 상승과 성층 강화가 가장 뚜렷하게 나타나는 해역으로, 이러한 물리적 변화는 독도 주변 연안 생태계에도 영향을 미칠 수 있다. 한반도 주변해역에서 수행한 장기관측 결과, 동해의 표층 수온 상승폭은 지난 35년간 2.04°C 정도를 나타내어, 서해(1.44°C)와 남해(1.27°C)보다 상대적으로 컸다(국립수산과학원 2025). 동해 표층과 수심 100 m 간의 수온차는 매년 0.01°C씩 증가경향을 나타내며, 성층화 특성이 강화되는 추세이다(국립수산과학원 2025). 태평양 순년 진동(Pacific Decadal Oscillation, PDO)과 동해의 해면 고도는 경년 변동성 규모에서 뚜렷한 상관관계가 보고되었다(박 2019). 태평양 순년 진동이 양의 위상일 때 동해 해수면이 낮아지며 이때 유입하는 난류가 약화된다고 알려져 있다(Gordon and Giulivi 2004; 박 2019; Shin et al. 2022). 동해의 표층혼합층 깊이는 계절과 기후변동 특성과 관련하여 뚜렷하게 차이를 나타내는데, 특히 여름철(6–8월)에는 강한 성층으로 인해 매우 얕은(10–20 m) 반면, 겨울철(12월–2월)에는 대기 냉각과 혼합의 강화로 매우 깊어지는(> 200 m) 것으로 보고된 바 있다(Trusenkova et al. 2005; Lim 2012). 이와 같은 해양환경 특성은 독도 주변 연안역에서도 동물플랑크톤 군집의 변동성을 유발할 수 있는 중요한 환경 요인으로 작용할 수 있다.
우리나라 동쪽 끝에 위치한 독도의 주변해역(독도를 기준으로 약 100 km 반경 범위)은 대마난류로부터 분지된 동한난류와 북한한류가 만나 형성된 전선과 소용돌이 구조의 영향을 받는 곳으로 알려져 있다(Chang et al. 2004; Teague et al. 2005; 해양수산부 1999). 독도 근접 연안역(독도 해안선으로부터 약 1 km 이내)은 바람과 해류의 상호작용으로 인해 수층 혼합이 활발하게 일어나며, 저층으로부터 유입된 영양분이 표층으로 유입되어 식물플랑크톤의 생산량을 증가시켜 동물플랑크톤 생물량이 풍부한 곳으로 알려져 있다(강 등 2002; Baek et al. 2018; Lee et al. 2021). 일차생산력에 영향을 미치는 환경요인은 온대해역에서 빛, 수온, 그리고 영양염류이고, 아열대와 열대해역에서는 유광층 아래에서부터 공급된 영양염류이다(Wafar et al. 1983; Behrenfeld and Falkowski 1997). 따라서, 일반적으로 섬 주변해역에서 해류의 흐름과 교차, 수직혼합과 함께 식물플랑크톤과 동물플랑크톤 생물량의 증가가 관찰되었다(강 등 2002; Baek et al. 2018; Lee et al. 2021). 이러한 물리·지형적 특성으로 인해 독도 연안역은 외해와 연안의 영향을 동시에 받는 해역으로 생물학적 변동성이 크게 나타날 수 있다.
독도 연안역의 동물플랑크톤 군집에 대해서는 그동안 일부 계절 조사 및 외해 중심의 연구가 수행되어 왔다. 독도 연안역에서 부유생태계를 구성하는 다양한 주요 구성성분 가운데 동물플랑크톤을 대상으로 한 연구 사례로는 수직분포와 주야수직이동(박과 최 1997), 독도 주변에서 춘계와 추계의 종조성과 개체수(강 등 2002)와 2006년부터 2015년까지 독도 인근해역에서 군집의 변동 특성(강 등 2016)이 있다. 이 중에 독도의 연안역에서 조사된 사례는 강 등(2002)의 연구가 유일하다. 독도 연안역은 섬으로부터 배출되는 물질의 직접적인 영향을 받는 동시에 외양역으로부터의 영향이 함께 작용하는 곳이다. 이 곳에서 동물플랑크톤 군집의 구조와 변동을 규명하는 것은, 독도 연안역 생태계의 기초 생산 및 상위 영양단계로의 에너지 전달 가능성의 이해를 위해 필요하며, 특히 환경요인과의 단·장기적 관계 이해에 중요하다. 그러나 최근접 연안역을 대상으로 다년간 계절 조사를 통해 중형동물플랑크톤 군집의 변동 특성을 분석한 연구는 제한적인 실정이다.
본 연구는 독도 연안역에서 중형동물플랑크톤 군집의 개체수와 조성을 3년간 계절적으로 채집 및 분석하여, 환경과 기후변동 관점에서 연계성을 이해하고자 하였다. 세부적으로, (1) 독도 최근접 연안역에서 출현하는 중형동물플랑크톤의 종 조성과 개체수의 시·공간적 특성 분석, (2) 계절적 군집특성과 우점종 변화 파악, (3) 환경요인과 군집 간의 상관관계를 분석하여 군집변동의 주요 요인을 도출하고자 하였다. 이를 위해 3개년에 걸쳐 수행된 계절조사를 바탕으로 계절성의 연간차이를 종합적으로 분석하였으며, 그 결과를 통해 독도 연안역 생태계에서 동물플랑크톤 군집특성을 단기 및 장기적 관점에서 이해하고, 기후변화와의 연관성을 이해하기 위한 기초자료를 제공하였다.
2. 재료 및 방법
조사 해역
본 연구는 2018년부터 2020년까지 4계절(봄, 여름, 가을, 겨울)에 독도 최근접 연안역 5개 정점(정점 A–E)에서 총 12회 조사하였다(Fig. 1). 현장조사는 한국해양과학기술원 연구선 온누리호와 이어도호에 승선하여 환경요인(수온, 염분, 엽록소-a)을 측정하고 동물플랑크톤을 채집하였다.
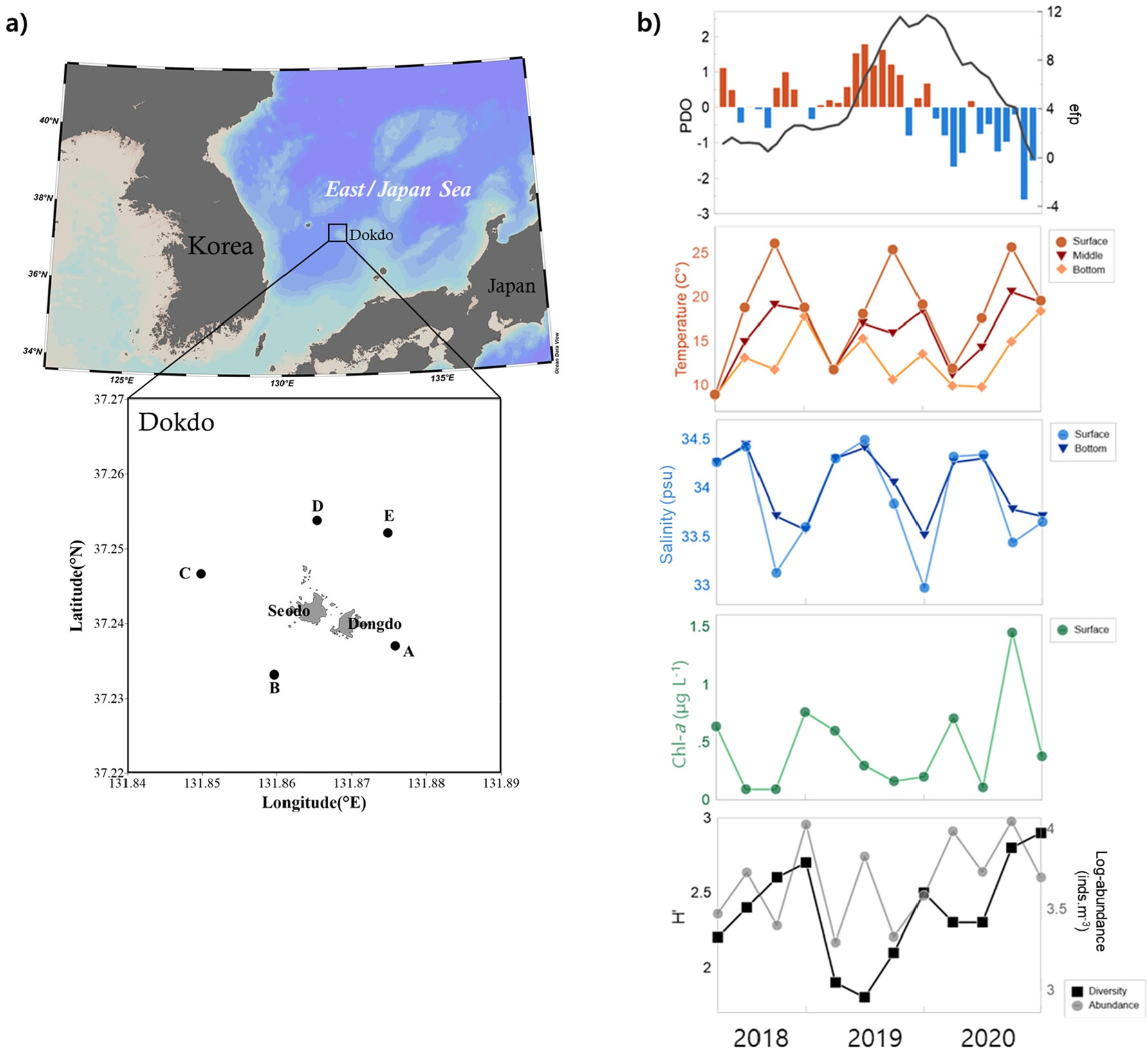
Fig. 1.
a) Map of sampling stations in the coastal waters of Dokdo, East Sea. b) Interannual variability (2018-2020) of hydroclimatic factors and environmental profiles: Pacific Decadal Oscillation (PDO), seawater temperature, salinity, chlorophyll-a concentrations, total abundance (log-transformed, inds. m-3) and species diversity index of mesozooplankton. Structural changes are shown by the cumulative sum of ordinary least square residuals (OLS-based CUSUM process denoted by the black thick line in the PDO phase
기후요인
Pacific Decadal Oscillation (PDO: 태평양 순년 진동) 지수는 미국 국립해양대기청(National Oceanic and Atmospheric Administration, NOAA)의 National centers for environmental information (https://www.ncei.noaa.gov/access/monitoring/pdo/, 접속일자 2025.06.15.)의 자료를 이용하였다. 태평양 순년 진동은 북태평양 해역의 해수면 온도 변동성에 대한 주요 경험 직교 함수(Empirical Orthogonal Function, EOF)를 기반으로 계산된 값으로서(Mantua 1999), 중위도 태평양에 영향을 미치는 대표적 장기 기후 변동성 패턴이다. 북위 20도 이북, 알래스카에서 캘리포니아에 이르는 동태평양 지역의 대륙 표면 기온에 큰 영향을 미치며, ENSO (El Niño-Southern Oscillation, 엘니뇨-남방진동)와 동시에 밀접하게 연관된 상호작용으로 한반도에 영향을 미친다고 알려져 있다(Park 2019; Kim et al. 2014).
현장 환경요인 측정
현장 수온과 염분은 SBE 911 CTD (Sea-Bird Electronics Inc.)을 이용하여 수직분포를 측정하였다. 표층 엽록소-a (Chlorophyll-a) 분석을 위한 해수 시료는 CTD 로제트 샘플러에 장착된 Niskin 채수병(General Oceanics)으로 채수하여, 유리섬유 필터(Whatman GF/F)에 낮은압력(125 mmHg 이하) 에서 여과 후 초저온 냉동고에(-80°C) 보관하였다. 이후 여과지를 90% 아세톤에 담아 냉장 암조건(4°C)에서 24시간 동안 엽록소를 추출한 후, 형광광도계(10-AU, Turner Designs)에서 엽록소-a 농도를 측정하였다(Parsons et al. 1984).
중형동물플랑크톤 채집과 분석
동물플랑크톤의 채집수심을 결정하기 위해 수온과 염분의 수직분포를 토대로 수온약층(thermocline depth)의 상부와 그 이하에서 해저면의 상부 약 5 m까지의 두 층을 구분하였다. 중형동물플랑크톤의 층별채집은 정선상태에서 폐쇄네트(망구직경 60 cm, 망목크기 200 µm) (General Oceanics)로 목표수심까지 네트를 내린 후, 30 m/min에서 50 m/min의 속도로 표층까지 수직예인하였다. 네트에 여과된 해수량은 네트 입구에 부착된 유량계(HydroBios backrun stop)의 회전수로 계산하였다. 코드엔드 버킷에 모인 채집시료를 1 L 폴리에틸렌 시료병에 옮겨 담은 후 중성 포르말린을 첨가하여 최종 농도가 약 5%가 되도록 고정한 후 실험실에 보관하였다.
실험실에서 시료병 내 중형동물플랑크톤 시료를 균등하게 혼합 후, 파이펫(10 mL)으로 5–10 mL의 부분시료(subsample)을 채취하여 계수판(Bogorov counting tray)에서 동정 및 계수하였다. 요각류는 종 수준까지, 그 외 중형동물플랑크톤은 속(genus) 수준 또는 분류군(taxa)별로 현미경(Zeiss Model Discovery SteREO V8, Germany)하에서 동정하였다. 계수된 중형동물플랑크톤 시료는 단위체적 당 개체수(inds. m-3)로 최종 환산하였다. 본 연구에서는 계절 및 경년 변동 특성의 일관된 비교를 위해, 수온약층을 기준으로 구분하여 채집된 상·하 수층의 자료를 합산한 전체 수층의 통합된 자료를 사용하여 분석하였다.
Pyrocystis pseudonoctiluca는 주로 열대와 아열대에 분포하는 원생생물로서, 평균 세포 크기는 약 330 µm 이다(Rivkin et al. 1982). 야광충(Noctiluca scintillans)의 크기와 형태와 흡사한 위 생물이 동물플랑크톤 네트에 대량으로 채집되어 형태와 크기를 고려하여 중형동물플랑크톤 군집에 우선 포함하여 계수하였다.
통계분석
중형동물플랑크톤 군집분석은 정점별 출현한 중형동물플랑크톤의 종조성과 개체수를 바탕으로 Bray-Curtis 지수를 산출한 후, 유사도 지수(similarity index)에 기반한 덴드로그램(dendrogram)으로 시각화하였다. 각 그룹의 구분에 영향을 미치는 중형동물플랑크톤 종들의 기여도를 파악하기 위해 SIMPER (similarity percentage) 분석을 하였으며, 자료 분석은 Primer version 6 (Primer-E)를 이용하였다. 군집분석 결과에서 도출된 그룹별 우점종과 환경요인 간의 관계는 정준상관분석(Canonical Correspondence Analysis, CCA)법으로 분석했고, R 프로그램으로 수행했다. 생물 다양성 지수와 환경조건과의 관계는, Pearson 상관분석(Pearson’s product-moment correlation)과 부트스트랩 리샘플링 기법(Casini et al. 2009)으로 수행하였다. 본 분석에서는 각 시계열 데이터를 대상으로 2,000회 반복적인 무작위 재추출을 수행하여 상관계수의 신뢰 구간을 산출하고 통계적 유의성을 검증하였다. 시각화에는 Grapher version 25 (Golden Software, LLC)와 Sigmaplot version 14.5 (SYSTAT) 소프트웨어를 활용하였다.
3. 결 과
기후요인
동해에 영향을 미치는 주요 기후와 환경요인의 계절별 변동성을 나타냈다(Fig. 1). 태평양 순년 진동은 연구 기간 중 양의 위상에서 음의 위상으로 전환되었다. 2019년 겨울까지 양의 위상이 지속되다가 11월과 12월이 지나면서 음의 위상으로 전환되어, 북서태평양에서 고온 현상이 우세할 것으로 나타났다. 연근해 해역의 환경과 기후 패턴에서 유의미한 구조적 변화가 관측되었으며, 특히 겨울철 수온 상승과 봄·여름의 빈번한 강우로 인해 염분이 감소한 것으로 판단된다.
독도 환경요인
수온은 모든 수층에서 계절 변동이 뚜렷했는데, 특히 표층에서는 여름철(평균 25.7°C) 고수온이 관측되었고 다른 계절에 비해 표층과 저층 사이의 온도차가 가장 컸다(Fig. 1b). 수층 간의 수온 차이는 2019년 여름철에는 표층이 중층보다 약 9.4°C가 높았던 반면, 2020년 여름철에는 그 차이가 좁아져서 약 5.1°C였다. 염분은 표층에서 저층까지 33.0–34.5 psu의 범위(평균 34.0 psu)였고 계절 변동이 뚜렷하였다. 가을철에 염분이 가장 낮았고(평균 33.4), 여름철에 수층 간의 염분 차이가 0.5 psu로 나타나 성층화가 강화된 양상을 보였다. 표층 엽록소-a 농도는 0.1–1.5 µg L-1범위(평균 0.5 µg L-1)였고, 2020년 여름철에 가장 높았다(Fig. 1b).
계절별 중형동물플랑크톤 군집특성
3년간의 조사 기간 동안 중형동물플랑크톤 군집은 계절별로 일정한 경향을 보였으나, 동일한 계절이라 하더라도 연도에 따라 총 개체수와 우점 분류군 조성에는 뚜렷한 차이가 나타났다. 이러한 연도별 변동을 개괄적으로 비교하기 위해 일부 결과는 평균값으로 제시하였다. 중형동물플랑크톤의 총 개체수는 뚜렷한 계절성과 연간변동을 나타냈는데, 특히 2018년 가을철과 2020년 여름철에 개체수가 가장 높았다(Fig. 2a). 2018년 가을철에는 요각류 성체와 미성숙체(73%)가 우점했고, 2020년 여름철에는 야광충의 형태와 크기가(평균 330 µm) 유사한 Pyrocystis pseudonoctiluca가 우점하고(63%), 방산충류가 차우점하였다(29%)(Fig. 2a).
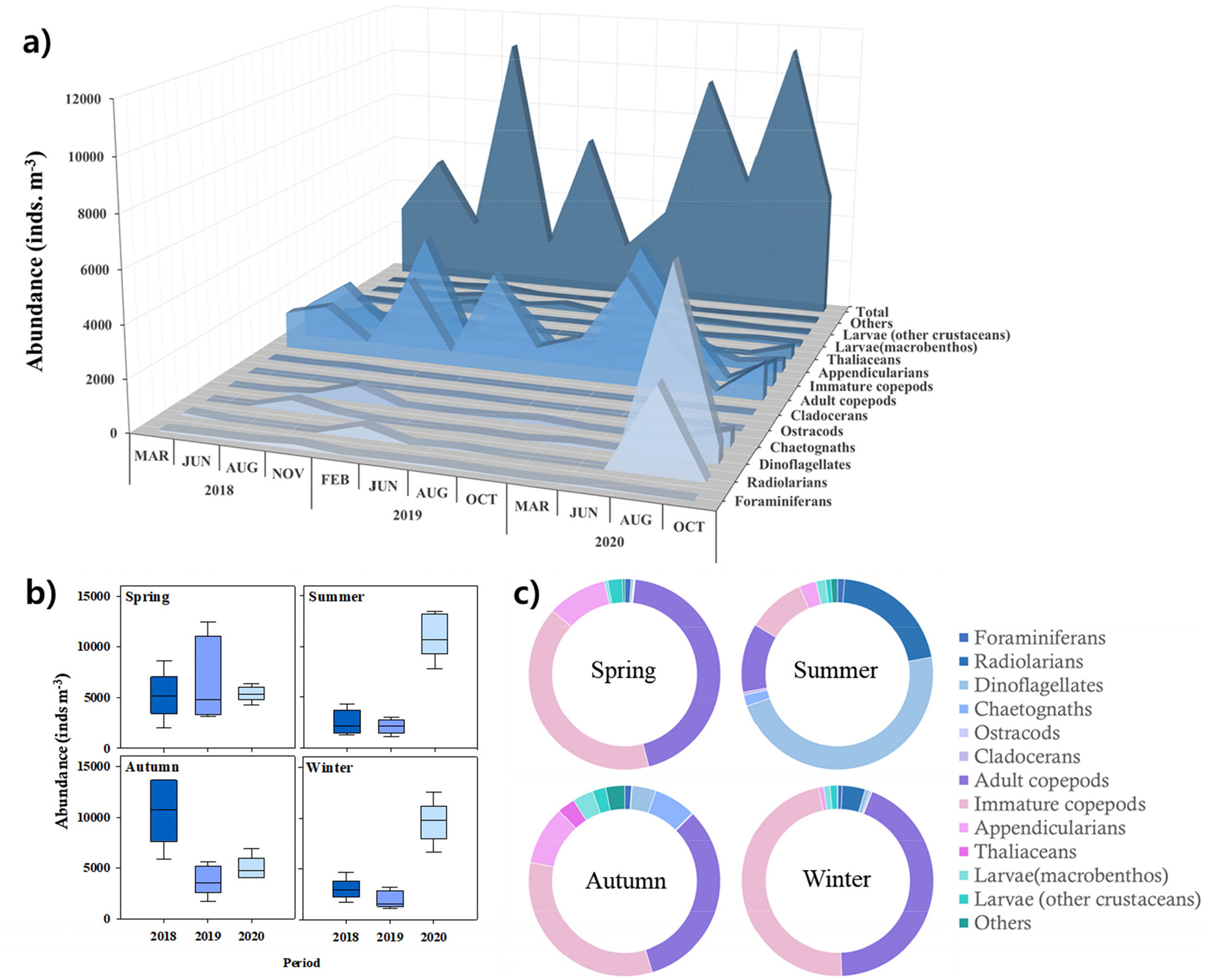
주목할 부분으로서, 2020년 여름과 겨울에 관찰된 평균 동물플랑크톤 개체수가 2019년에 비해 약 5배, 2018년 대비로도 약 3배 높아 가장 큰 증가폭을 보였다(Fig. 2b). 그 차이는 2019년 12월 이후 태평양 순년 진동 지수가 양에서 음으로 바뀌는 시기와 엽록소-a 농도가 증가하는 경향과 함께 나타났다(Figs. 1b and 2b). 계절적으로 중형동물플랑크톤 군집구조의 차이는 뚜렷했는데, 겨울철과 봄철에 요각류 중심(평균 88%, 성체와 미성숙체)의 특성을 나타낸 반면, 여름철에는 원생생물(48%)과 방산충류(21%)의 점유율이 높았고, 가을철에는 요각류(66%), 유형류(10%)와 모악류(7%)의 점유율이 다른 계절에 비해 약 3배 높았다(Fig. 2c).
계절별 중형동물플랑크톤 군집 변화와 환경요인 간의 관계 분석
조사기간 동안 출현한 중형동물플랑크톤 군집을 분석한 결과, 유사도 52% 기준으로 주요 6개 그룹으로 구분되었다(Fig. 3). 2018년 겨울철(그룹 A), 여름철(그룹 E)과 가을철(그룹 F)의 경우 각 한 개의 그룹으로 구분되었고, 그 외 모든 해의 봄철과 2020년 겨울철이 한 그룹으로(그룹 C), 2020년 여름철과 가을철이 한 그룹으로(그룹 B), 2019년 겨울철, 여름철과 가을철이 한 그룹으로 구분되었다(그룹 D). 그룹 B와 E의 경우, P. pseudonoctiluca 주도하였고, 나머지 그룹들은 긴노요각목(Calanoida) 미성숙체 또는 Paracalanus 미성숙체가 주도하였다(Fig. 3, Table 1). SIMPER (similarity percentage)분석에서 총 기여율 합이 51% 이상인 분류군을 확인한 결과, 차 우점종인 요각류 Oncaea clevei, Oithona 미성숙체, Paracalanus 미성숙체, Paracalanus parvus s.l.과 유형류 Fritillaria spp.와 Oikopleura spp.의 점유율로 인해 계절 특이적 차이를 나타냈다(Fig 3, Table 1). 그룹 C에서 태평양 순년 진동 지수가 2019년 봄철(1.2)을 제외하고 모두 음의 값을 나타냈는데, 이때 긴노요각목 요각류인 Paracalanus 미성숙체, Paracalanus parvus s.l., P. aculeatus, Scolecithrix 미성숙체와 검물벼룩목(Cyclopoida)의 Oithona 미성숙체와 O. similis가 주도하였다.
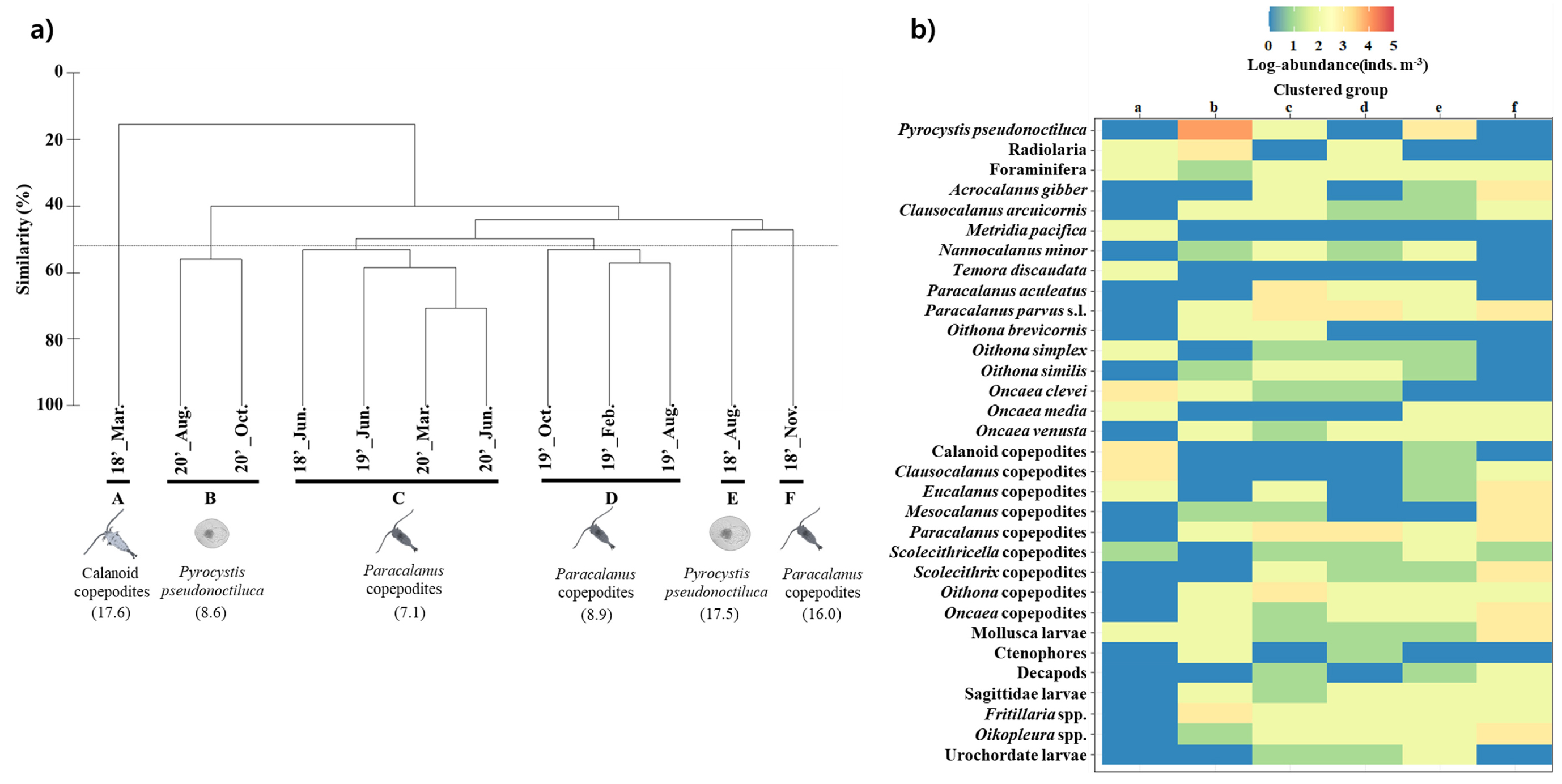
Table 1.
Composition of mesozooplankton taxa identified by SIMPER analysis based on cluster analysis (Bray-Curtis similarity)
정준상관분석
정준상관분석을 통해 독도 연안역 중형동물플랑크톤 종과 환경요인 간의 관계를 분석한 결과, 제 1축과 제 2축은 각각 44.9%와 20.36%의 분산을 설명했다. 표층 수온과 빗해파리류(Ctenophores)와, 저층 수온과 유형류 Fritillaria spp.가, 태평양 순년 진동과 중층 수온과 요각류 Eucalanus 미성숙체와, 엽록소-a 농도와 요각류 Paracalanus 미성숙체와 모악류 유생과 유의한 양의 상관관계를 나타냈다(Fig. 4). 태평양 순년 진동이 음의 위상일 때(-1.1) 서태평양의 수온은 상승하는데, 2019년에 비해 2020년 겨울철 Eucalanus 미성숙체의 개체수가 증가했다(133 inds. m-3). 이는 계절풍 약화 및 혼합층 변화와 연계된 먹이 환경의 변동이 미성숙체의 생존과 성장에 영향을 주었을 가능성을 시사한다.
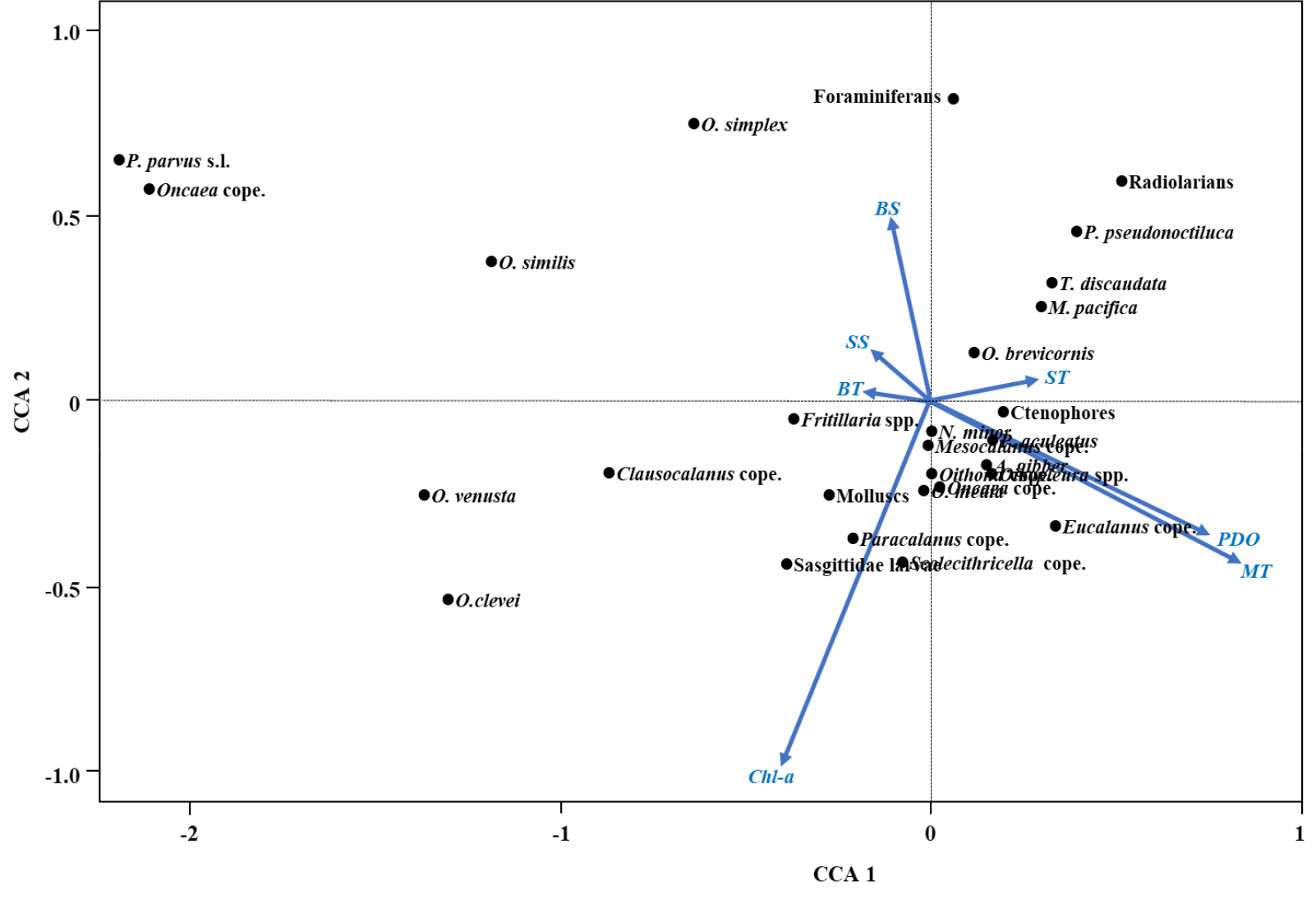
Fig. 4.
Canonical Correlation Analysis (CCA) biplot with first two axes from CCA of Dominant mesozooplankton taxa (black points) and environmental variables (blue vectors) including surface temperature (ST), middle layer temperature (MT), bottom layer temperature (BT), surface salinity (SS), bottom layer salinity (BS), surface chlorophyll-a concentrations (Chl-a), Pacific Decadal Oscillation (PDO). Dominant mesozooplankton includes foraminiferans, Radiolarians, Pyrocystis pseudonoctiluca (P. pseudonoctiluca), Ctenophores, Sagittidae larvae, Acrocalanus gibber (A. gibber), Clausocalanus arcuicornis (C. arcuicornis), Metridia pacifica (M. pacifica), Nannocalanus minor (N. minor), Paracalanus aculeatus (P. aculeatus), Paracalanus parvus s.l. (P. parvus s.l.), Temora discaudata (T. discaudata), Oithona brevicornis (O. brevicornis), Oithona similis (O. similis), Oithona simplex (O. simplex), Oncaea clevei (O. clevei), Oncaea media (O. media), Oncaea venusta (O. venusta), Clausocalanus copepodites (Clausocalnaus cope.), Eucalanus copepodites (Eucalanus cope.), Mesocalanus copepodites (Mesocalanus cope.), Paracalanus copepodites (Paracalanus cope.), Scolecithricella copepodites (Scolecithricella cope.), Scolecithrix copepodites (Scolecitrix cope.), Oithona copepodites (Oithona cope.), Oncaea copepodites (Oncaea cope.), Oikopleura spp., Fritillaria spp. and molluscs
4. 고 찰
계절별 동물플랑크톤 군집특성
독도 연안역 중형동물플랑크톤 군집은 계절에 따라 뚜렷한 우점 분류군의 전환이 나타났으며, 이는 수온과 먹이 환경의 계절적 변동에 대한 생태적 반응으로 해석될 수 있다. 동물플랑크톤 군집의 계절별 특성은, 겨울철과 봄철에 요각류 성체와 미성숙체의 점유율이 매우 높아(평균 88%) 요각류 중심의 군집조성을 보였고, 특히 Paracalanus와 Oithona 미성숙체, P. parvus s.l.와 O. clevei의 점유율이 높았다. P. parvus s.l.는 온대와 열대해역에 분포하며(Cornils and Held 2014), 연간 국내 연안역에 출현하는 우점종 중 하나이며 수온 범위10–22°C에서 출현한다고 알려져 있다(박과 최 1997; 강 등 2002; 문 등 2010). P. parvus s.l.는 남해에서 여름철에 가장 낮고 가을철에 가장 높게 출현하였고(문 등 2010), 봄철과 가을철에 우점하며, 특히 남해 전선역에서 가장 높은 점유율을 차지했고(Kim et al. 2019), 남해 연안수의 영향이 강하게 미치는 곳에서 점유율이 특히 높았다(장 등 2012). 동해 전선역 부근에서 상대적으로 높은 수온(18–21°C)에서 출현하였고 난류수 선호종으로 알려져있다(박과 최 1997). 북부 캘리포니아 해류의 영향을 받는 연안에서는 온대종으로 알려진 P. parvus s.l.가 외해역과 상관관계가 가장 높았는데, 이는 온대 지시종으로서 환경 변화를 관측하는데 유용할 수 있음을 의미한다(Dumelle et al. 2021).
요각류 중에서도 Paracalanus parvus 종복합체는 온대 연안역에서 환경 변동에 민감하게 반응하는 대표적인 분류군으로 알려져 있다. 본 연구에서 Paracalanus 미성숙체의 개체수와 엽록소-a 농도의 상관관계가 유의하게 나타났다(r = 0.65, p < 0.05). P. parvus의 월간 난생산율(egg production rate, EPR)은 봄철과 여름철에 높고 수온보다 엽록소-a 농도와 상관성이 높았으며, 특히 엽록소-a 농도는 유생(nauplii) 성장에 긍정적인 영향이 있다(Kang and Kim 2023). 요각류 출현 개체수는 여름철에 비해 봄철에 높았으며, 이는 봄철에 비교적 높은 엽록소-a 농도와 밀접하다고 알려져 있다(서 등 2018). 본 연구에서 엽록소-a 농도가 2020년 여름철에 가장 높았는데, 일반적으로 독도 주변해역에서는 동한난류가 이동한 경로주변에서 높은 엽록소-a 농도(5 mg m-3)가 관측되었고 침편조류 Heterosigma akashiwo가 우점하였다(백과 김 2018). 2020년 여름철의 경우 독도 인근 외해역에서도 비교적 높은 엽록소-a 농도 (0.6 µg L-1)가 관측되었고 이때 강한 서수도 수송량과 동한난류로 인해 동해연안역의 생산성이 독도 인근해역까지 전달되었을 수도 있음을 시사한다(Kim et al. 2024).
독도 인근 해역의 동물플랑크톤 군집 변동에 대해서는 이미 일부 선행 연구가 보고된 바 있다. Lee et al. (2019)는 독도 주변 해역에서 요각류를 중심으로 한 동물플랑크톤 군집이 계절 및 수온 변화에 따라 뚜렷한 변동을 보인다고 보고하였으며, Pardianto et al. (2019) 역시 독도 인근 해역에서 동물플랑크톤 군집 구조가 환경 요인과 밀접하게 연관되어 있음을 제시하였다. 본 연구에서 관찰된 계절별 우점 분류군의 변화와 경년 규모의 변동 특성은 이러한 선행 연구 결과와 전반적으로 일치하는 경향을 보이며, 장기간의 반복 조사 자료를 통해 독도 연안역 동물플랑크톤 군집의 변동 특성을 보다 구체적으로 확장한 사례로 볼 수 있다.
여름철에 관찰된 원생생물의 상대적 증가는 수층 성층 강화와 고수온 조건에서 나타나는 부유생물 군집의 전형적인 반응으로 이해될 수 있다. 여름철에는 비교적 높은 개체수의 야광충과 P. pseudonoctiluca가 포함된 원생생물(48%)과 방산충류의 점유율(21%)이 높았다. 야광충은 적조원인종으로서 온대해역에서 열대해역까지 분포하며(Fonda-Umani et al. 2004), 독도 주변해역에서 2014년 봄철에 야광충 대발생이 관측된 바 있다(강 등 2016; 강 2020). 야광충 개체수의 주요 증가요인은 높은농도의 먹이생물과 해양학적 과정(용승, 소용돌이, 해류, 바람 등)등이 있는데(강 2020), 독도 인근 외해역에서는 수온과 야광충 개체수가 같이 증가한 사례가 있다(Kim et al. 2024). 2018년부터 독도 주변해역에서 관측된 P. pseudonoctiluca는 2020년도 여름철과 가을철(군집분석, 그룹 B)에 가장 높았는데 이는 발광성 와편모조류이며 주로 아열대와 열대해역에서 분포하는 종이며, 운동성이 없고 오직 부력을 통해 수직이동을 하며 수온이 20°C 이상인 해역에서 출현한다(Rivkin et al. 1982; Hauslage et al. 2017). 이러한 결과는 2020년에 나타난 태평양 순년 진동의 음의 위상으로 알 수 있듯이 강한 서수도 수송량을 통해 대량 유입되고 해양열파의 발생으로 인해 개체수가 유지되었을 가능성을 시사한다(Kim et al. 2024).
젤라틴성 동물플랑크톤의 증가는 계절적으로 변화하는 먹이 입자 크기와 수층 혼합 특성을 반영하는 생태적 지표로 활용될 수 있다. 가을철의 요각류, 유형류와 모악류의 출현율이 다른 계절에 비해 높았다(Fig. 2). 유형류는 미세한 유기입자(박테리아, 식물플랑크톤 등)를 여과섭식하며 형태적으로 mucous house를 주기적으로 생성하고 폐기하는 과정을 통해 유기물질 수송에 크게 기여를 하는 젤란틴성 플랑크톤이다(Alldredge 1981; Capitanio et al 2018). 유형류 Oikopleura spp.는 난류수 선호종이며 동해 가을철(11월)에 표층과 수심 20 m의 범위에서 우점하였고(박과 최 1997), 남해에서는 겨울철에 우점하며 밀도와 염분과 양의 상관성을 나타냈다(이 등 2021). 2019년부터 2020년까지의 겨울철과 가을철에 태평양 순년 진동이 음의 위상(-0.56)일 때 Eucalanus 미성숙체의 개체수가 증가하는 경향(평균 258 inds. m-3)을 보였다. 이러한 증가는 동중국해와 황해에 난류의 북상과 난온수괴 확장에 따라 난온성 요각류인 Eucalanus subcrassus가 겨울철에도 분포하는 측면과(Wang et al. 2003) 관련이 있어 보인다. 다른 한편, 차가운 수괴 조건에서 계절 휴면(diapause) 또는 수직이동을 통해 개체수를 유지하는 생존 전략이 알려져 있다(Jackson and Smith 2016; Baumgartner and Tarrant 2017).
기후변동과 동물플랑크톤 특성
조사 기간 동안 관찰된 중형동물플랑크톤 군집의 경년 변동은 단순한 계절성뿐 아니라, 해당 시기의 해양환경 변동성과 함께 해석될 필요가 있다. 계절적 기후 조건의 변동성(Fig. 1)은 단순한 환경 배경을 넘어서, 독도 연안역의 동물플랑크톤 계절 변동을 해석할 때 핵심 단서로 활용될 수 있다. 특히 겨울철 수온 상승과 여름철 강우에 따른 염분 감소는 동물플랑크톤 군집 조성과 생활사 전략에 직·간접적으로 영향을 미쳤을 가능성이 있다. 본 연구 기간 중 태평양 순년 진동은 2019년 12월에 양에서 음의 위상으로 전환되었고, 이는 북서태평양의 해수온 상승과 연계될 수 있다(McAfee 2014). 태평양 순년 진동은 널리 북태평양 해양과 생태계에 연결된 기후 패턴으로, 해수면 온난화 패턴과 해양 열파 발생과 관련이 깊은 것으로 보고된 바 있다(Ren et al. 2023). 또한, 겨울철 수온상승과 봄·여름철의 빈번한 강우에 따른 염분 감소는 플랑크톤 군집의 조성, 영양염 분포, 그리고 먹이망 내 구성성분 간의 상호작용에 직·간접적으로 작용할 수 있으며, 이는 기존에 보고된 동해 연안 생태계 변동 양상과도 맥락을 같이한다(강 등 2002; Chang et al. 2004). 따라서 본 연구에서 확인된 물리환경 변화는 생태계 수준의 상향 효과(식물플랑크톤 생산력 강화)와 하향 효과(동물플랑크톤 생활사 조절)의 잠재적 기작으로 이해될 수 있다.
동아시아 연안해(East Asian Marginal Seas)의 해양 환경 변화는 수온, 성층과 수괴 이동 등 물리적인 요인에 의해 직접적으로 결정되며, 태평양 순년 진동은 이러한 장기 변동성을 반영하는 기후지수이다. 이 지수는 관측된 변동이 어떤 기후적 배경과 연계되어 있는지를 해석하는 데 활용되는 간접적 지표이다(Wang and Wu 2022). 양의 태평양 순년 진동 위상은 알류샨 저기압의 강화와 편서풍의 증대를 통해 북서태평양의 수온 저하를 유도하며(Huo et al. 2008), 해양학적 환경조건과 생태계 기능에 연쇄적인 영향을 미칠 수 있다. 예를 들어, 수온 상승과 염분 증가는 수층의 안정화를 강화하여 혼합을 억제하고, 영양염의 표층유입이 제한됨으로써 표층 엽록소-a 농도의 감소로 이어질 수 있다. 실제로 수온 및 염분의 증가, 엽록소-a 농도의 감소 등 뚜렷한 해양학적 변화는 여러 기존 연구에서 보고된 바 있으며(Kim et al. 2014; Jung et al. 2021), 이는 곧 식물플랑크톤 생산력의 저하와 더불어 동물플랑크톤 개체군과 생활사 전략에도 직·간접적인 영향을 미칠 수 있다(Lynam et al. 2017; Browning and Moore 2023). 따라서 이러한 기후학적 변화는 상향 효과(영양염과 일차생산력 조절)와 하향 효과(먹이망 구조 및 소비자 군집 변화)를 동시에 매개하는 중요한 배경기작으로 이해될 수 있다.
장기기후변동 지수 중 하나인 태평양 순년 진동을 통해 단기간에 관찰된 중형동물플랑크톤 군집의 변동이 계절 혹은 연간변이를 넘어 기후변동 관점에서도 이해가 될 수 있다. 동중국해에서 보고된 바와 같이, 이러한 기후 요인은 동물플랑크톤, 특히 요각류 Calanus sinicus와 같은 주요 종의 시간적 변동성과 관련이 있었다(Rebstock and Kang 2003; Molinero et al. 2018; Ma et al. 2019). 요각류 개체수 변동성이 태평양 순년 진동 기반 환경 변화와 맞물려 나타나는게 일반적이므로, 주요 요각류의 종 조성이 급격히 바뀐다면 부유생물 생태계 내 체제전환(regime shift)도 확인할 수 있다(Keister et al. 2011; Liu et al. 2015).
본 연구에서 계절에 따라 변화하는 환경요인(수온, 염분, 엽록소-a)과 중형동물플랑크톤 군집의 반응이 같은 계절 안에서 비슷한 방향으로 나타나는 경향을 확인하였다. 이와같은 계절 동시적 반응(coherent seasonal responses)은 환경 변화가 여러 분류군에 동시에 영향을 미칠 때 흔히 관찰되는 현상으로, 이는 해양과 담수 플랑크톤 군집에서도 보고된 바 있다(Honda et al. 2023; O’Brien et al. 2023). 특히, 2019년 이후 태평양 순년 진동 지수가 음의 위상으로 전환되는 시기에 표층 수온 상승과 엽록소-a 증가가 동시에 관측되었고, 이와 같은 환경조건 변화는 같은 계절 내에서 Paracalanus 미성숙체의 개체수 증가로 이어졌다. 표층 엽록소-a 농도와 Paracalanus 미성숙체 개체수 사이에 유의한 양의 관계가 확인되었는데, 이는 상향 효과를 통한 동시성 반응의 정량적 단서를 제공한다. 이러한 관측 결과는 태평양 순년 진동의 직접적인 조절 요인은 아니지만, 해양환경 변화의 배경적인 변동을 반영함으로써 독도 연안역의 생산성 변화와 중형동물플랑크톤 우점종 변화 또는 특정 분류군의 증가와 같은 변화에 간접적으로 연계될 수 있음을 시사한다. 본 연구는 독도 연안역 동물플랑크톤 군집의 계절적 구조와 단기 변동성을 제시하는 기준자료로서 의미를 가지며, 기후 인자의 연속적인 장기 변동 맥락 속에서 관찰될 수 있음을 보여준다. 이러한 결과는 향후 장기 관측 자료가 축적될 경우, 기후 변동과 해양 생태계 반응을 통합적으로 이해하는 데 기초 자료로 활용될 수 있다.
5. 결 론
본 연구에서 3년(2018년–2020년) 동안 계절별로 독도 최근접 연안역에서 중형동물플랑크톤 군집을 조사하여 계절과 경년 변동성을 분석하였다. 중형동물플랑크톤 군집은 뚜렷한 계절특성을 나타냈는데, 겨울철과 봄철에는 요각류, 특히 Paracalanus spp.와 Oithona spp.가 우점한 반면, 여름철에는 원생생물인 야광충과 P. pseudonoctiluca의 개체수가 증가하였고, 가을철에는 요각류와 더불어 젤라틴성 동물플랑크톤 분류군이 출현하였다. 특히 2019년 12월 이후 태평양 순년 진동이 양에서 음의 위상으로 전환되면서 독도 최근접 연안 해양생태계에서 엽록소-a 농도와 중형동물플랑크톤의 총 개체수가 동시에 증가하였다. 정준상관분석 결과, 주요 우점종은 수온, 엽록소-a 농도 그리고 태평양 순년 진동과 유의한 양의 상관관계를 보였고, 이는 기후 변동과 물리적 환경 변화가 중형동물플랑크톤 군집구조 변화와 관련이 높음을 시사한다. 본 연구는 독도 연안역의 환경요인 간의 복잡한 상호관계 속에서 나타난 동물플랑크톤의 시간적 변동이 기후변화 관점에서 이해될 수 있음을 제시하였다.
