

1. Introduction
Oncaeid copepods are relatively small marine cyclopoids that are distributed worldwide in high abundances (Huys and Boxshall 1991; Nishibe and Ikeda 2004). To date, the family Oncaeidae Giesbrecht, 1893 [“1892”] consists of 108 valid species in seven genera, with members distributed across the world’s oceans (Walter and Boxshall 2020). They are common prey of juvenile fish (Itoh et al. 2014) and play an important role in material cycling in marine ecosystems (Koski et al. 2017).
In Korea, ecologists started recording oncaeid copepods belonging this family at the end of the 20th century but have reported only four species: Oncaea media Giesbrecht, 1891, O. mediterranea (Claus, 1863), O. venusta Philippi, 1843, and Triconia conifera (Giesbrecht, 1891)[= Oncaea conifera] (Shim and Park 1982; Shim and Lee 1986; Kim et al. 1993; Park and Choi 1997; Park et al. 1998). Subsequently, several taxonomic studies on these copepods were conducted in Korean waters by Wi et al. (2008, 2009, 2010, 2011, 2012), who found four species of the genus Oncaea Philippi, 1843 and six species of the genus Triconia Böttger-Schnack, 1999. More recently, three species of Triconia were described from the Korea Strait by Cho et al. (2013), and one species, T. dentipes (Giesbrecht, 1891), was listed without a taxonomic description by Song et al. (2017). To date, 18 species belonging to two genera in the family Oncaeidae are known in Korea: seven species of Oncaea and 11 species of Triconia (NIBR 2019).
During an investigation of the zooplankton community in Korea waters, a previously unrecorded species, O. prendeli Shmeleva, 1966 was found in south sea of Jeju Island. Oncaea prendeli was originally described from the Adriatic Sea in the Mediterranean by Shmeleva (1966), but the original description lacked details such as the number of setae on the distal endopod segment of P1 and P4 as well as the caudal ramus. As the holotype of O. prendeli designated by Shmeleva does not exist anymore and the species needed to be reviewed, Huys and Böttger- Schnack (2007) redescribed the species and discussed dis-crepancies in detail based on the newly designated neotype.
The present study describes O. prendeli collected in south sea of Jeju Island and compares to the species from the Adriatic Sea in the Mediterranean described by Huys and Böttger-Schnack (2007). In addition, we compared spine lengths on the exopods and endopods of swimming legs 2–4 with those of closely related species, O. tregoubovi Shmeleva, 1968, and three species of the genus Spinoncaea Böttger-Schnack, 2003 newly established to accommodate with the species of Oncaeaivlevi-group.
2. Materials and Methods
The material examined in the present study was sampled using a fine-mesh conical zooplankton net (60 cm mouth diameter, 60 µm mesh) deployed from a depth of 110 m to the surface at station KS4 (32°36'46.66"N, 126°42'51. 89"E) in south sea of Jeju Island on October 7, 2008 (Fig. 1). Sample was immediately fixed with 99.9% ethyl alcohol on board. For the morphological examination, the specimen was dissected with tungsten needles and mounted in fluoromount-G on an H-S slide, which then was sealed with transparent nail varnish. All figures were prepared using a drawing tube attached to a differential interference contrast microscope (Leica DM2500, Germany). Scale bars in the figures are given in micrometers (μm).
The terminology for body and appendage morphology follows that of Huys et al. (1996), using the following abbreviations: Al, antennule; A2, antenna; ae, aesthetasc; exp, exopod; enp, endopod; exp (enp)-1 (2, 3) to denote the proximal (middle, distal) segment of a three-segmented ramus; P1–P6, first to sixth thoracopod.
3. Description
Family Oncaeidae Giesbrecht, 1893 [“1892”]
Genus Oncaea Philippi, 1843
Oncaea prendeli Shmeleva, 1966
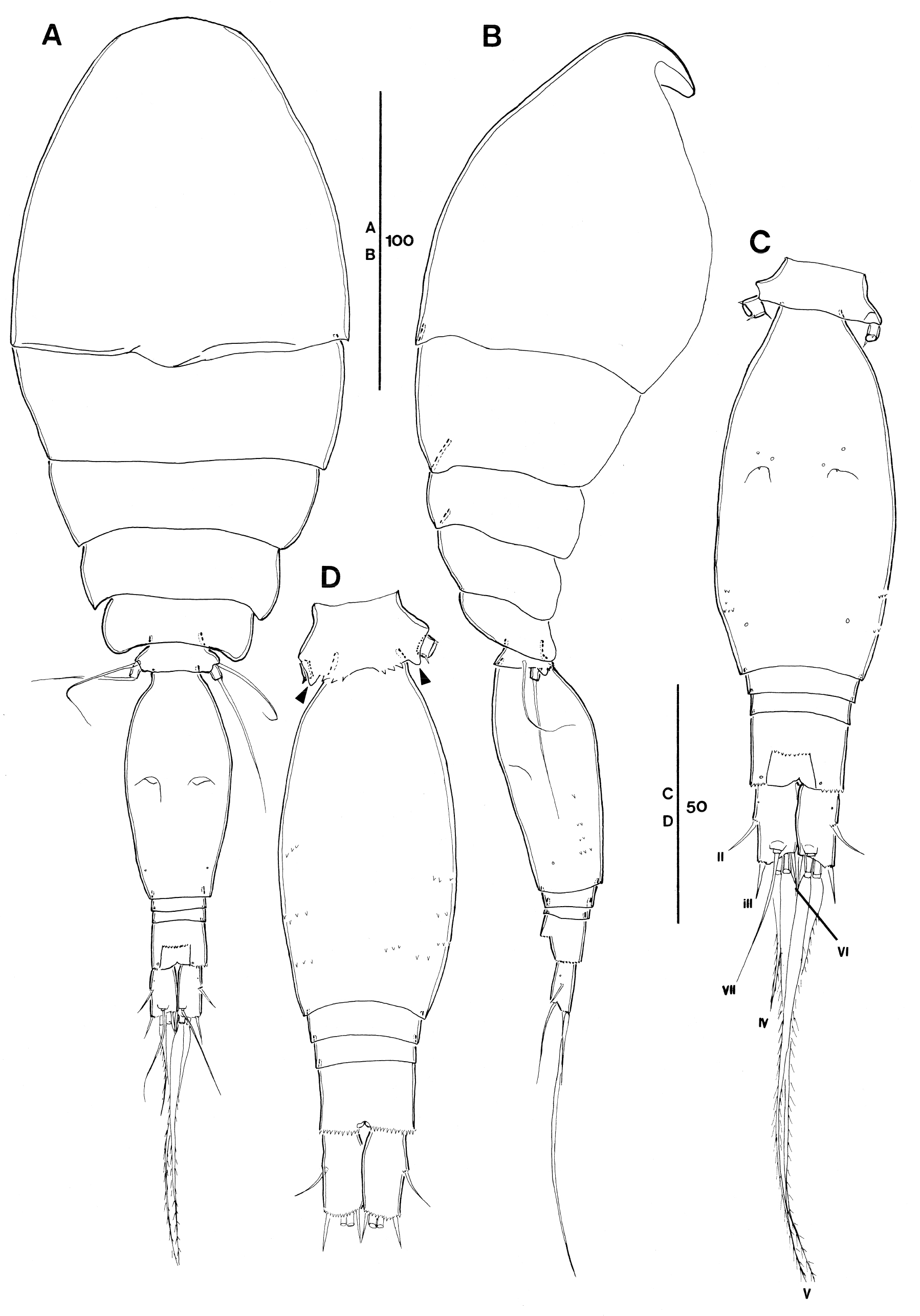
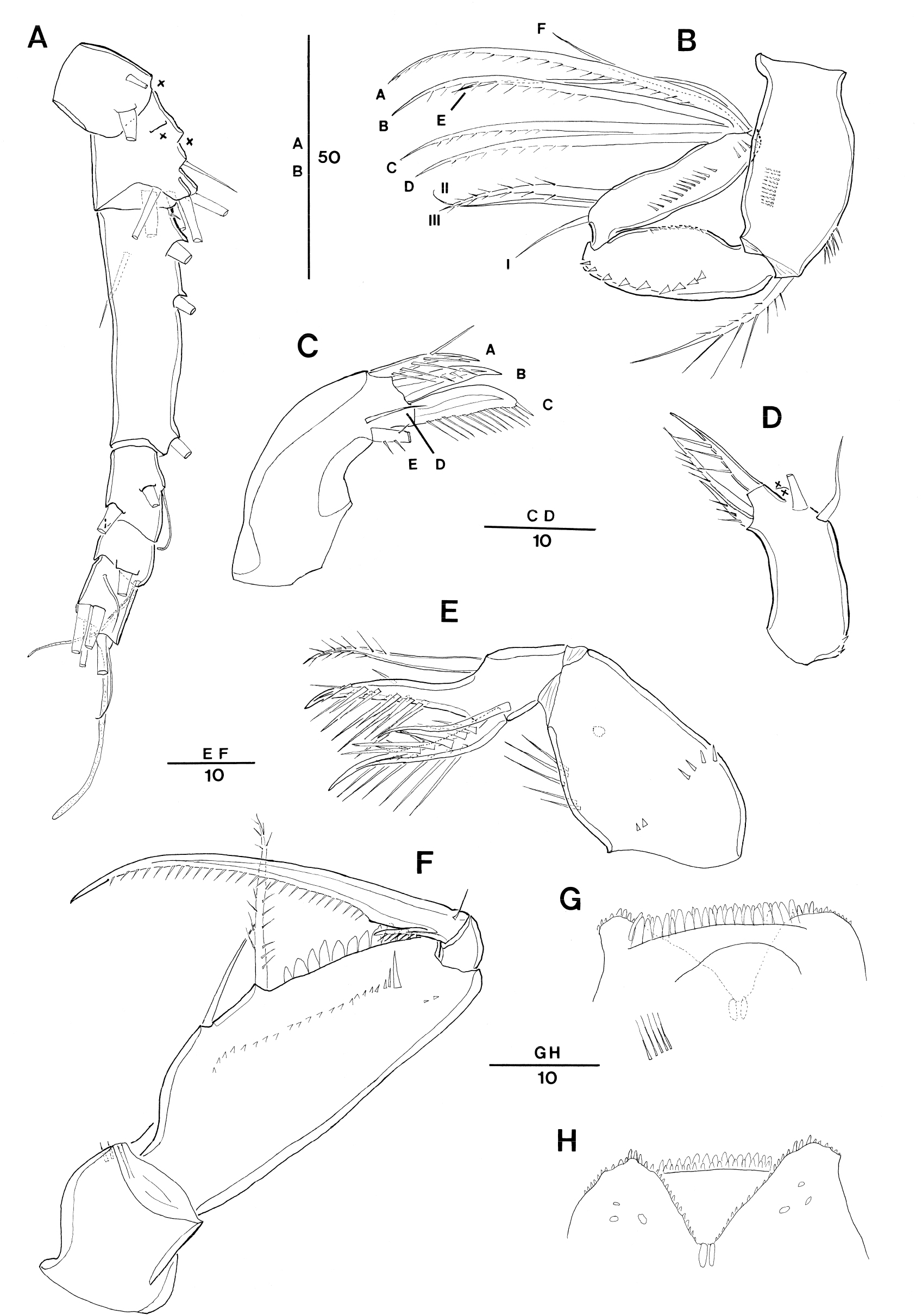
Fig. 3
Oncaea prendeli Shmeleva, 1966. Female (A) antennule (crosses indicate a seta being lost in the specimen examined whose position was inferred from the insertion points on the exoskeleton); (B) antenna, distal elements on distal endopod segment numbered using capital letters, lateral elements indicated by Roman numerals; (C) mandible, individual elements indicated by capital letters (E element damaged in the specimen); (D) maxillule (elements on outer lobe damaged, crosses indicate elements being lost in the specimen); (E) maxilla; (F) maxilliped, anterior; (G) labrum, anterior (long setules on the left side damaged); (H) labrum, posterior
Oncaea prendeli Shmeleva, 1966, 933–934, Figs. 2.1–2.9 (♀), 2.10–2.11 (♂); Shmeleva, 1969, 8–10, 27, Figs. 5a–i (♀), 6a–i (♂); Huys and Böttger-Schnack, 2007, 95–111, Figs. 2–6.
Type locality: Adriatic Sea; Stn. Lokrum; 42°38.5'N, 18°02.0'E; 25–50 m
Material examined: One female dissected and mounted on an H-S slide. This specimen was collected in south sea of Jeju Island, Korea, St. KS4 (32°36'46.66"N, 126°42'51. 89"E); 0–110 m depth; 7 October, 2008.
Female. Body (Fig. 2A and B) length: 325 μm (based on one specimen).
Exoskeleton weakly chitinized. Prosome about 2.1 times as long as urosome, excluding caudal rami, about 1.8 times urosome length including caudal rami. P2-bearing somite without conspicuous dorsoposterior projection in lateral aspect (Fig. 2B). Pleural areas of P4-bearing somite with rounded posterolateral corners.
P5-bearing somite with 3 pairs of midventral spinous processes and 1 pair of ventrolateral lobate process (arrowed in Fig. 2D).
Genital double-somite [this compressed by coverslip on an H-S slide] oval-elongate, about 2.1 times as long as maximum width (measured in dorsal aspect) and 2.8 times as long as 3 postgenital urosomites combined (measured in dorsal aspect); maximum width measured at about halfway the length. Paired genital apertures located dorsally at halfway the genital double-somite length; armature represented by 1 small spinule. Secretory pore location on dorsal surface as in Fig. 2C; ventrolateral surface with several rows of minute spinules (Fig. 2B–D). Anal somite slightly wider (about 1.2 times) than long; about as long as caudal rami (Fig. 2C). Secretory pore discernible on both sides of anal opening posteriorly. Anterior margin of anal opening with transverse row of denticles. Posterior margin of somite serrates ventrally and laterally (Fig. 2C and D).
Caudal ramus (Fig. 2C) 1.8 times as long as wide. Armature consisting of 6 elements: setae II and III small and slender, spinform, and unornamented, seta II slightly longer than seta III; seta IV more than 4 times as long as seta III, moderately lanceolate, and bipinnate at distal part; seta V about 2.8 times as long as seta IV, lanceolate; seta VI about as long as seta III, unornamented; dorsal seta VII arising from small peduncle, unornamented, and biarticulate at base. Dorsolateral anterior surface (Fig. 2C) with secretory pore anteriorly. Inner margin of caudal ramus unornamented. Few spinules present on base of insertions of setae II and III. Posterior margin of ramus ornamented with minute spinules ventrally (Fig. 2D).
Antennule 6-segmented (Fig. 3A). Armature formula: 1-[3], 2-[8], 3-[5], 4-[2? (one seta not discernible) + ae], 5-[2 + ae], 6-[5 + (1 + ae)]. Setation pattern very difficult to discern, one specimen examined with incomplete setation pattern.
Antenna 3-segmented, distinctly reflexed (Fig. 3B). Coxobasis with row of spinules along outer and inner margins; with a long seta at inner distal corner, ornamented bilaterally with spinules of varying length and short spinules. Endopod 2-segmented, segments elongate, of equal length; distal segment distinctly slender than proximal one. Proximal endopodal segment oval, outer margin bearing short spinular row; posterior surface with row of short, strong denticles along inner margin. Distal endopodal segment about 4 times as long as wide, with narrow cylindrical base articulating with proximal segment; posterior surface with row of tiny spinules along outer margin; lateral armature consisting of 1 bipinnate spiniform seta (III) and 2 naked setae (I, II), and seta I shortest; distal armature consisting of 4 long spiniform setae (A–D), ornamented with spinules unilaterally along entire (A) or half (C–D), 2 naked setae (E, F), seta E longer than seta F, and posterior seta G absent.
Labrum (Fig. 3G and H) indistinctly bilobed (damaged, some anterior part not figured). Each lobe with 3 marginal teeth on distal margin and row of short spinules or denticles along outer ventral and inner margin. Median concavity covered anteriorly by overlapping rows of hayaline spatulate spinules, anterior surface with paired row of long setules (damaged, only one side figured in fig. 3G). Posterior wall of medial concavity with 2 chitinized spinous teeth, flanked by row of tiny denticles or spinules. Posterior surface with 3 pairs of secretory pores located distally on each lobe.
Mandible (Fig. 3C) without surface ornamentation on gnathobase; with 5 clearly separated elements: 3 setae and 2 blades. Ventral seta (A) as long as ventral blade (B), with long setules along dorsal side; ventral blade (B) strong and spiniform, with row of setules on posterior side; dorsal blade (C) strong and broad, spinulose along entire dorsal margin; seta D short and naked; seta E (damaged) setiform and bipinnate.
Maxillule (Fig. 3D) (damaged, some elements not figured). indistinctly bilobed, with few spinules around proximal outer margin. Inner lobe (praecoxal arthrite) subcylindrical, with 3 elements: outermost element spiniform, swollen at base, ornamented with row of spinules; others setiform and sparsely unipinnate. Outer lobe with 4 elements (of which 2 elements missed as indicate crosses in the Fig. 3D).
Maxilla (Fig. 3E) 2-segmented, comprising syncoxa and allobasis. Syncoxa unarmed, ornamented with 2 spinular rows and 1 secretory pore on surface; inner margin with row of long spinules. Allobasis produced distally into slightly curved claw bearing 2 rows of strong spinules along the inner margin and single spinule on outer margin; outer margin with strong bipinnate seta extending to tip of allobasal claw; inner margin with slender naked seta and strong basally swollen spine with 2 rows of very long spinules along the inner margin and few spinules along outer margin.
Maxilliped (Fig. 3F) 4-segmented, comprising syncoxa, basis and 2-segmented endopod. Syncoxa unarmed, surface ornamented with few spinules subdistally. Basis oval-elongate, and with 2 spiniform setae, the distal one being about 1.4 times the length of proximal one, with 2 rows of short spinules along entire length and the proximal one naked; fringe of short spatulate spinules along palmar margin between distal seta and articulation with endopod; anterior surface with row of short spinules and row of spinules of varying length near articulation with endopod as illustrated in Fig. 3F. Proximal endopodal segment small and unarmed. Distal endopodal segment drawn out into long curved claw, with spinnules along entire concave margin; accessory armature consisting of short, slender seta on outer proximal margin and unipectinate spine basally fused to inner proximal corner of claw.
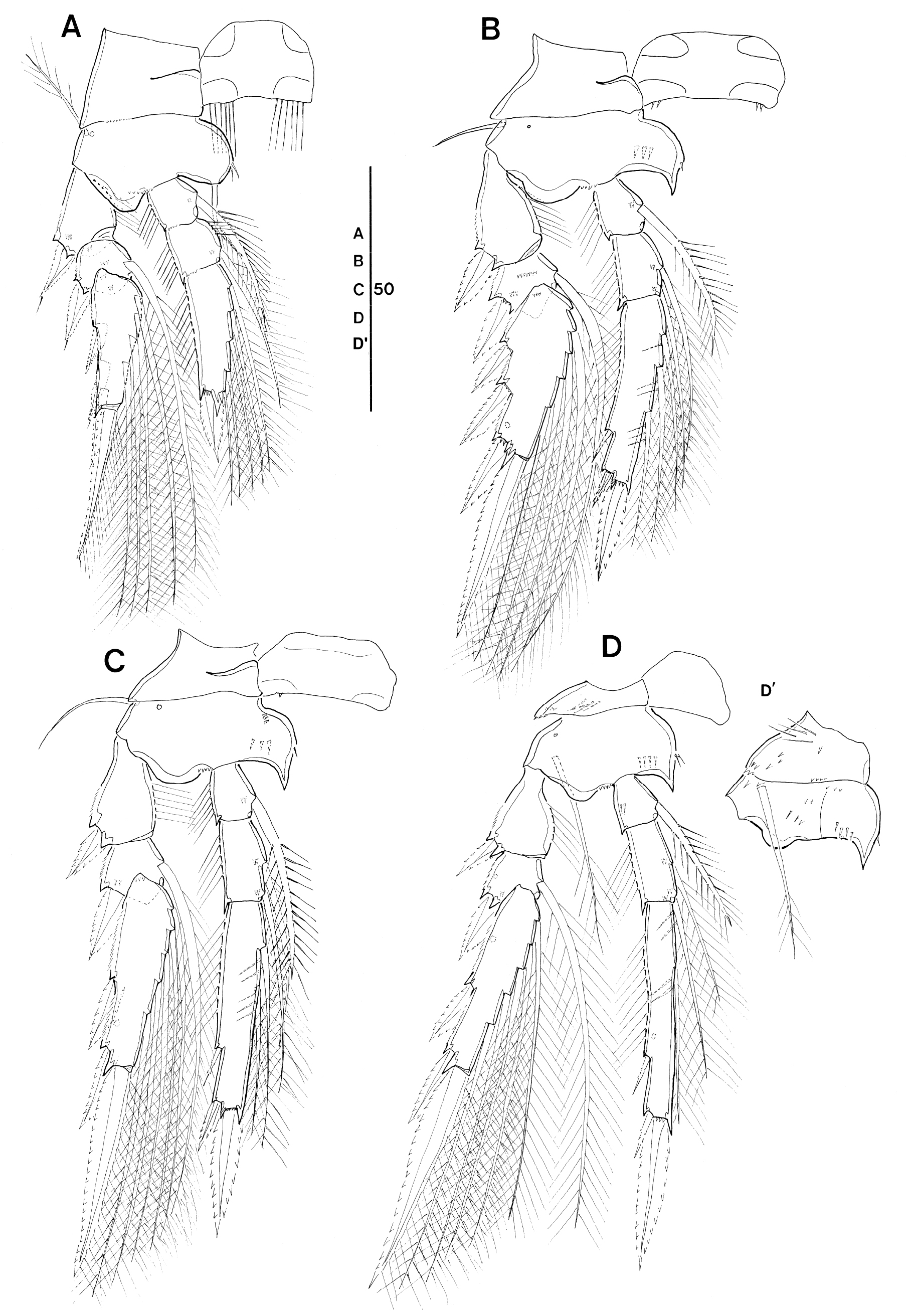
Swimming legs 1–4 biramous (Fig. 4A–D), with 3- segmented rami. Intercoxal sclerites well developed; with paired patches of very long setules along ventral margin in P1 (Fig. 4A), sparsely ornamented in P2 and P3 (Fig. 4B and C), and unornamented in P4 (Fig. 4D). Coxae and bases with surface ornamentation as shown in Fig. 4A–D, coxa of P4 with patch of long, fine setules on posterior surface (Fig. 4D'). Bases with short and sparsely plumose (P1) or short and naked (P2 and P3) or long, and sparsely plumose (P4) outer seta; with secretory pore at outer proximal corner of anterior surface; inner portion produced adaxially into rounded (P1) or pointed (P2–P4) process, bearing short spinule(s) along inner margin. Inner basal seta on P1 naked. Respective legs without distinct length differences between exopod and endopod.
Leg armature formula See Table 1.
Table 1.
Leg armature formula
Exopods. Outer margins of exopodal segments with serrated hyaline lamella; inner margin of proximal exopod segments with long setules. Exopodal segments with surface ornamentation as shown in Fig. 4A–D and with secretory pore located on posterior surface of distal segments. Hyaline lamellae on outer spines moderately developed; outer and terminal spines of P1 with subapical tubular extension, except for proximalmost spine on distal exopod segment. Distal (terminal) spines slightly longer than distal exopod segments. Length ratios of exopodal spines of P2–P4 are given in Table 2.
Table 2.
Comparison of ratios of exopodal spine length to endopodal spine length for females of Oncaea prendeli, O. tregoubovi, and three Spinoncaea species
| O. prendeli | O. tregoubovi | S. ivlevi | S. humesi | S. tenuis | |||
| Korean waters | Mediterranean Sea | Red Sea | |||||
| L ratio spines P2 enp-3 | OSDS:DS | 0.39/0.35 | 0.38 | 0.42 | |||
| ODS:DS | 0.43/0.49 | 0.52 | 0.55 | 0.59 | 0.56 | 0.50 | |
| L ratio spines P3 enp-3 | OSDS:DS | 0.34/0.37 | 0.44 | 0.35 | 0.59 | 0.41 | 0.37 |
| ODS:DS | 0.42/0.45 | 0.52 | 0.45 | 0.54 | 0.48 | 0.41 | |
| L ratio spines P4 enp-3 | OSDS:DS | 0.27/0.33 | 0.34 | 0.30 | 0.38 | 0.35 | 0.30 |
| ODS:DS | 0.33/0.33 | 0.38 | 0.37 | 0.46 | 0.41 | 0.28 | |
| L ratio exopodal spines on P2 | OS exp-1: OS exp-2 | 0.89/0.89 | 0.92 | 1.31 | 1.67 | 1.20 | 1.26 |
| OS exp-1: 1st OS exp-3 | 1.09/1.14 | 1.00 | 1.70 | 1.43 | 1.04 | 1.33 | |
| L ratio exopodal spines on P3 | OS exp-1: OS exp-2 | 0.80/0.78 | 0.74 | 1.14 | 1.29 | 1.20 | 1.12 |
| OS exp-1: 1st OS of exp-3 | 0.75/0.80 | 0.67 | 1.27 | 1.00 | 1.14 | 0.79 | |
| OS exp-1: 2nd OS exp-3 | 0.67/0.68 | 0.65 | 0.97 | 0.93 | 1.00 | 0.73 | |
| L ratio exopodal spines on P4 | OS exp-1: OS exp-2 | 0.74/0.76 | 0.74 | 0.87 | 1.24 | 1.18 | 1.11 |
| OS exp-1: OS exp-3 | 0.75/0.80 | 0.77 | 0.87 | 1.11 | 1.18 | 0.91 | |
| OS exp-1: 2nd OS exp-3 | 0.64/0.65 | 0.63 | 0.76 | 1.00 | 0.91 | 0.77 | |
| Reference | This study* | Huys and Böttger-Schnack (2007)** | Böttger-Schnack (2003)** | ||||
Abbreviations: P, thoracopod; DS, distal spine; L, length; ODS, outer distal spine; OS, outer spine; OSDS, outer subdistal spine
**Values from re-examination of figures in Huys and Böttger-Schnack (2007) (Figs. 4 and 8) and Böttger-Schnack (2003) (Figs. 4, 10, and 14)
Endopods. Outer margins of endopodal segments with fringe of long setules, except for proximal endopod segment of P4; inner margin of P1 enp-2 with 4 long spinules. Inner setae of proximal endopod segment slightly swollen and ornamented with spinules bilaterally. Inner setae of P4 enp-1 and enp-2 slightly reduced in length. Endopodal segments with surface ornamentation as shown in Fig. 4A–D and with secretory pore on posterior surface in P4 enp-3. Inner margin of P1 enp-3 terminating in long, bi-modal process obscuring insertion of distalmost inner seta. Distal margins of P2–P3 enp-3 not produced into conical process, but apical pore of reduced process discernible, located laterally between outer distal spine and distal spine. Outer subdistal spines on P2–P4 enp-3 naked, slightly shorter than outer distal spine; length ratios of outer subdistal spine (OSDS) and outer distal spine (ODS) relative to distal spine (DS) given in Table 2. Inner setae of P2–P4 enp-3 with spinule comb along proximal inner margin; this comb also present on distal inner seta of P4 enp-2.
P5 (Fig. 2A and B) comprising long outer basal seta, which is naked, reaching as far as genital apertures [this seta gets lost during handling] and a small free segment representing the exopod. Exopod slightly longer than wide (Fig. 2A), ornamented with small spinule ventrally and small spinous process dorso-laterally at posterior margin (Fig. 2C) and armed with single long, naked seta, reaching as far as 2/3 from anterior margin of genital double-somite.
P6 (Fig. 2C) represented by operculum closing off each genital aperture; armed with a short spinule.
4. Discussion
The female O. prendeli has the following morphological characteristics: (1) elongated and slender distal endopod segment on antenna; (2) armature of the antenna with six distal setae without seta G, and three lateral setae without seta IV; (3) presence of two medial teeth on the posterior surface of the concavity on labrum; (4) ornamentation on the female maxilliped with spatulate spinules; (5) presence of midventral spinous processes on the P5-bearing somite; (6) ornamentation along the ventral margin of the P1 intercoxal sclerite, with a paired row of long, fine setules; (7) presence of a spinular row along the inner margin of P1 enp-2; (8) reduction of the P5 exopod with a single seta; and (9) lanceolate form and length of caudal seta IV.
The O. prendeli collected from south sea of Jeju Island shares the aforementioned characteristics with the Mediterranean specimens redescribed by Huys and Böttger- Schnack (2007). However, the Korean specimen appears a few minor morphological differences: (1) in the antenna, the length-to-width ratio of the distal endopod segment is slightly larger (ratio = 4) than that of the Mediterranean specimens (ratio = 3); (2) the ornamentation on the coxobasal seta on the antenna is bilateral (vs. unilateral in the Mediterranean specimens); (3) the maxilla presents long spinules along the inner margin of the syncoxa in the Korean specimen (vs. absent in the Mediterranean specimens); and (4) caudal seta IV is about two-fifths of seta V and approximately four times as long as seta III (vs. approximately a half of seta V and five times longer than seta III in the Mediterranean specimens. In addition, the fourth segment of the antennule has only two setae in the Korean specimen, whereas in the Mediterranean specimens it has three setae. Furthermore, the fifth segment of the antennule carries an aesthetasc in the Korean specimen, but it is absent in the Mediterranean specimens. These discrepancies in micro-characteristics may necessitate another review of O. prendeli based on morphological and/or molecular information of specimens collected globally from various oceans regions including Korean waters.
Oncaea prendeli is most closely related to O. tregoubovi and three Spinoncaea species, based on recently re- examined phylogenetic relationships (Böttger-Schnack 2003), but it can be clearly distinguished from those four species based on the absence of seta G on the distal endopodal segment of the antenna as mentioned in characteristic (2). Besides, O. prendeli has spatulate spinules on the maxilliped, whereas O. tregoubovi does not, and caudal seta IV in O. prendeli is less than half or half the length of seta V, whereas it is approximately two-thirds of the length of seta V in O. tregoubovi. Among the aforementioned characteristics of O. prendeli, differences between O. prendeli and the Spinoncaea species are evident based on characteristics (6) and (9); i.e., except for S. tenuis Böttger-Schnack, 2003, the other Spinoncaea species do not have characteristic (6) (Böttger-Schnack 2003). Regarding characteristic (9), caudal seta IV in the Spinoncaea species is never more than three times as long as seta III (Huys and Böttger-Schnack 2007; their Table 3). In addition, the Spinoncaea species have only one outer spine in the distal endopod of P2, but O. prendeli possesses two spines.
Subtle morphological differences and/or small ornamentation details are significance for identification of oncaeid species (e.g. Böttger-Schnack 2003; Wi et al. 2010). To easily discriminate among related and sister species of the oncaeid group, which are very small and thus, have micro-characters that are difficult to analyze, proportional spine lengths on the exopods and endopods on P2–P4 were measured as previously described (Heron and Bradford-Grieve 1995; Böttger-Schnack 2005; Cho et al. 2017, 2019). The outer spine on proximal exopod segment relative to the outer spine on middle exopod segment of O. prendeli tends to be smaller than those in both O. tregoubovi and the Spinoncaea species, whereas differences in endopodal spine lengths among these five species are not informative (Table 2). Although such information is currently lacking, it is necessary to consider not only the relative lengths of the caudal setae but also the proportional outer spine lengths of the exopods in P2–P4 to clearly distinguish O. prendeli from closely related species.
The current distribution range of O. prendeli extends from in the Mediterranean Sea to the equatorial Indian Ocean (Huys and Böttger-Schnack 2007) and Tosa Bay in southern Japan. However, the latter Japanese record was based on an ecological study, not a taxonomic one (Nishibe et al. 2009). Herein, we report the occurrence O. prendeli in Korean waters. Thus, this species likely has a wide distribution range.
The species diversity of oncaeid copepods collected using a fine-mesh net (< 100 µm) is higher than those using other collection methods. Nishibe et al. (2009) added more than 20 species to those previously known when they used a 63 µm mesh net for their investigations. Cho (2011) also identified 19 previously unreported oncaeid species in the northeast equatorial Pacific after sampling with a 60 μm mesh net. Therefore, to discover new or unrecorded species of small oncaeids in Korean waters, it is necessary to use a fine-mesh net (mesh size ~60 µm).
