

1. Introduction
2. Material and Methods
3. Result and Discussion
Trigonium cf. curvatum A.Nair, R.Mohan, S.Shetye, S.Gazi & S.A.Jafar 2015
Amphitetras antediluviana Ehrenberg 1840
Halamphora pseudoholsatica (T.Nagumo & H.Kobayasi) J.G.Štěpánek & Kociolek 2018
Halamphora sydowii (Cholnoky) Levkov 2009
Luticola sp.
Tryblionella coarctata f. densestriata Álvarez-Blanco & S.Blanco 2014
Entomoneis japonica (Cleve) K.Osada 2002
4. Conclusion
1. Introduction
Submarine groundwater discharge (SGD) is increasingly recognized as a significant hydrological process through whic h terrestrial groundwater enters the coastal ocean, bypassing surface runoff pathways. SGD is defined as the flow of groundwater through permeable sediments or rock fractures directly into the marine environment, delivering not only freshwater but also substantial amounts of nutrients, trace metals, and dissolved gases to coastal systems (Kim et al. 2012; Lecher and Mackey 2018). Although SGD is estimated to represent approximately 10% of the global river discharge volume, its biogeochemical influence on coastal ecosystems is disproportionately significant due to the elevated concentrations of solutes it frequently transports (Taniguchi et al. 2002; Moosdorf and Oehler 2017).
The volcanic island of Jeju, situated south of the Korean Peninsula, serves as an exemplary natural laboratory for the study of SGD. Characterized by its basaltic geology, high permeability, and the absence of major rivers, Jeju Island contains over 900 documented groundwater springs, approximately 145 of which are located along the coastline and identified as SGD discharge sites (Hahn et al. 1997; Lee et al. 2004). Historically, these coastal springs have served as essential freshwater sources for the island's inhabitants and function as transitional zones where terrestrial and marine biogeochemical processes converge (Hwang et al. 2010; Moosdorf and Oehler 2017).
A notable characteristic of SGD in Jeju is the presence of highly mineralized, saline groundwater, commonly termed Jeju lava seawater. This groundwater, originating from deep volcanic aquifers, is characterized by substantially elevated concentrations of dissolved silica—reported to be up to 20 times higher than those found in typical coastal seawater—alongside appropriate levels of nitrogen, phosphorus, iron, and trace elements (Kim and Kim 2011; An et al. 2024). The geochemical distinctiveness of Jeju lava seawater is attributed to prolonged interactions between water and basaltic substrates. These conditions may create highly favorable environments for siliceous microalgae, particularly diatoms, which require silica for the formation of their frustules (Martin-Jézéquel et al. 2000). Nevertheless, the physicochemical properties of SGD systems can vary considerably depending on temporal and hydrological factors, resulting in a broad spectrum of salinity regimes ranging from freshwater-dominated to saline groundwater conditions.
SGD environments are characterized by intrinsic dynamism, exhibiting spatiotemporal variability in salinity, temperature, redox conditions, and nutrient concentrations due to the mixing of fresh groundwater and seawater (Ahmadi Musaabad et al. 2019; Lecher and Mackey 2018). These environmental gradients promote niche heterogeneity and are increasingly recognized as hotspots of biological diversity and functional complexity. Moreover, the mixing of water columns driven by SGD can facilitate the resuspension of benthic microalgae into the photic zone, thereby enhancing planktonic-benthic coupling (Lecher and Mackey 2018). Such environments have been associated with the proliferation of opportunistic macroalgae, such as Ulva, and the occurrence of phytoplankton blooms under nutrient-enriched conditions (Liefer et al. 2009; Kwon et al. 2017). Additionally, these gradients are anticipated to influence benthic diatom assemblages, which are highly sensitive to variations in salinity, nutrient availability, and substrate characteristics.
Diatoms (Bacillariophyta) represent a crucial group of primary producers in aquatic ecosystems, contributing significantly to the global cycling of carbon and silica. The species composition of diatoms is highly sensitive to environmental conditions, making them reliable indicators of water quality and ecosystem health (Srivastava et al. 2016; Serôdio and Lavaud 2022). Although previous studies have characterized diatom assemblages in the river mouths and estuarine zones of Jeju (Lee et al. 2001; Jung et al. 2014; Kim et al. 2019), no study has specifically investigated diatom communities associated with SGD springs in Korea. In particular, the floristic composition of diatoms in these groundwater-influenced microhabitats remains largely unexplored.
Floristic surveys of diatoms conducted in diverse regions worldwide, especially within alpine and karstic spring ecosystems, have revealed a high degree of species richness and ecological specificity. These studies often identify taxa adapted to narrow physicochemical niches (Żelazna-Wieczorek and Ziułkiewicz 2009; Cantonati et al. 2012). This body of research highlights the potential of springs, including those affected by SGD, to support unique microbial communities that are frequently underrepresented in conventional assessments of aquatic biodiversity.
This study examines the diatom flora of Chunggulmul, a coastal SGD spring located on the northeastern shore of Jeju Island, with particular emphasis on species not previously recorded in South Korea. The primary objective is to present a floristic account of diatoms from the Chunggulmul SGD spring. Morphological characteristics are documented through light microscopy (LM) and scanning electron microscopy (SEM) observations, and preliminary ecological context is provided for this groundwater-influenced coastal habitat.
2. Material and Methods
For the purposes of this study, samples were collected from the site designated as SGD (33º33'30.3''N, 126º45'4.3''E), referred to as Chunggulmul on Jeju Island, on July 5, 2021 (Fig. 1). Temperature and salinity were measured using a MP556 Multiprobe System (YSI, Yellow Springs, OH, USA) and recorded as 16.6°C and 2 PSU, respectively. The water samples were subsequently transferred into 50 mL conical tubes and preserved with 5% Lugol’s solution (Sigma Aldrich, St. Louis, MO, USA). A pretreatment procedure was implemented to investigate the fine structure and specific morphological characteristics of diatoms by removing organic matter from the samples. To eliminate organic material, samples were treated with nitric acid following the rapid cleaning method described by Trobajo and Mann (2019). A small aliquot of the sample was placed on a coverslip and air-dried, after which concentrated 70% nitric acid (HNO3) was added and heated on a hotplate at approximately 90°C. The acid was allowed to evaporate (approximately 10–15 min), and the treatment was repeated two to three times until no visible organic residue remained. The cleaned material was subsequently rinsed multiple times with distilled water to remove residual salts and air-dried prior to mounting. Finally, the cleaned samples were mounted in Naphrax (Brunel Microscopes, Chippenham, UK) and analyzed using an Eclipse Ni light microscope (Nikon, Tokyo, Japan) equipped with a ×100 oil immersion objective lens and a DS-RI2 digital camera (Nikon).
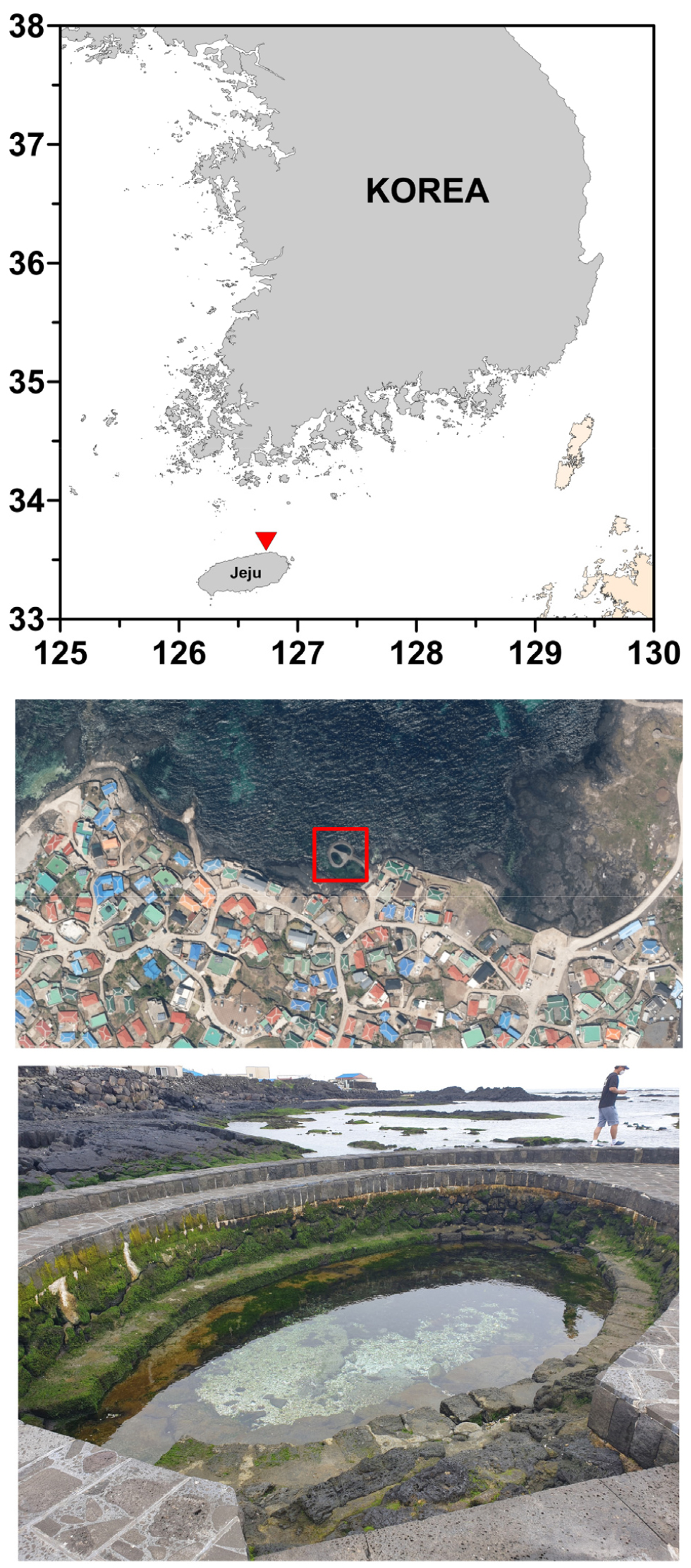
Fig. 1.
Study area and sampling site at Chunggulmul, Jeju Island. (a) Regional map showing the position of Jeju Island off the southern coast of the Korean Peninsula (red triangle). (b) Satellite image of Gimnyeong Village on the north-eastern shore of Jeju; the red box marks the oval, stone-lined coastal spring (Chunggulmul) investigated in this study. (c) Ground-level view of Chunggulmul. The satellite image included in this figure was sourced from Google Earth Map
For the examination conducted using a scanning electron microscope (SEM), the sample was initially fixed in a 5% Lugol's solution and subsequently filtered through a polycarbonate membrane with a pore size of 3 µm and a diameter of 25 mm (Advantec, Tokyo, Japan). The membrane was then rinsed with distilled water. Dehydration of the specimen was performed by sequential immersion in ethanol solutions of increasing concentration (10% to 100%), beginning at 10%. After dehydration through the ethanol series, the specimen was air-dried, and subsequently desiccated using tetramethylsilane (Sigma Aldrich). The membrane was mounted on a stub and coated with gold via sputter coating. SEM observations were conducted using a Sigma 500-VP field emission SEM (Zeiss, Göttingen, Germany) at an accelerating voltage of 2.0 kV and a working distance of approximately 5.4 mm, employing a secondary electron detector (SE2).
Diatom identification was conducted through consultation of various literature sources that provide comprehensive species descriptions of the species. Morphological observations focused on valve characteristics, including valve outline, raphe system, and striae pattern. Measurements, such as valve length, width, and striae density, were obtained from digital images; striae density was quantified by counting the number of striae within a 10 µm interval. For certain taxa, particularly rare or newly recorded species, only a limited number of individuals were available due to low abundance. Nevertheless, identification was based on consistent diagnostic morphological features observed in all available specimens using both LM and SEM, in accordance with standard taxonomic literature for each genus and species.
All images presented in this study were obtained from a total of three sources: two permanent slides prepared for LM and one SEM stub. All examined materials, including the permanent slides and SEM stubs, are archived in the National Marine Biodiversity Institute of Korea (MABIK) and are accessible for verification upon request. Additionally, the raw image data are also stored in an institutional repository.
The status of taxa as new records for Korea was determined by comparing the identified taxa with previously published Korean diatom records and the most recent national species checklists. These included the National Species List of Korea (NIBR 2025), provided by the National Institute of Biological Resources under the Ministry of Climate, Energy and Environment, and the National List of Marine Species (MABIK 2025), provided by the National Marine Biodiversity Institute of Korea under the Ministry of Oceans and Fisheries. Additionally, a review of previously published Korean diatom literature was conducted to verify the absence of prior records.
3. Result and Discussion
As presented in Table 1, a total of 48 diatom taxa were identified from discharge samples collected at the Chunggulmul spring. These taxa are classified into 28 genera, 25 families, 19 orders, and 3 classes, following the taxonomic framework established by AlgaeBase (Guiry and Guiry 2025). Among these, five species—Amphitetras antediluviana, Entomoneis japonica, Halamphora pseudoholsatica, Halamphora sydowii, and Tryblionella coarctata f. densestriata—represent new records for Korea. In addition, two taxa, Trigonium cf. curvatum and Luticola sp., could not be confidently assigned to known species and are therefore excluded from the list of new records. The status of these taxa as novel records was verified through comparison with previously published Korean diatom literature and the most recent national species checklists. These taxa are indicated with an asterisk (*) in Table 1. This section details the floristic composition and taxonomic accounts of the observed diatom assemblage, highlighting primary morphological features that differentiate them from closely related species.
Table 1.
Diatom taxa recorded from the Chunggulmul submarine groundwater discharge site on Jeju Island. Taxonomic classification, figure numbers, habitats, and key references are provided. Asterisks denote species-level taxa newly recorded in Korea. 'Sal.' refers to salinity preference, with the following classifications: M = Marine, B = Brackish, F = Freshwater, and S = Soil. Additionally, 'Ecol.' denotes ecological type, which includes PL = Planktonic, BE = Benthic, EP = Epipelic, EI = Epiphytic, and TP = Tychoplanktonic
| Classification | Fig. no | Habitat | Key References | |
| Sal. | Ecol. | |||
| Class Coscinodiscophyceae Round & R.M.Crawford 1990 | ||||
| Order Melosirales R.M.Crawford 1990 | ||||
| Family Melosiraceae Kützing 1844 | ||||
| Melosira inflexa (Roth) Guiry 2019 | 2–4, 73, 74 | M/B | TP | Witkowski et al. (2000); Hoppenrath et al. (2009); Joh (2010) |
| Melosira nummuloides C.Agardh 1824 | 5, 6, 75 | F/B | TP, EP | Witkowski et al. (2000); Hoppenrath et al. (2009); Joh (2010) |
| Order Paraliales R.M.Crawford 1990 | ||||
| Family Paraliaceae Kützing 1844 | ||||
| Paralia guyana MacGillivary 2015 | 7, 76 | M/B | TP | MacGillivary and Kaczmarska (2015); Yun et al. (2016) |
| Order Arachnoidiscales Round 1990 | ||||
| Family Arachnoidiscaceae Round 1990 | ||||
| Arachnoidiscus ehrenbergii Bailey 1849 |
9, 10, 77, 78 | M | EP, EI | Shim (1994) |
| Order Stephanopyxales Nikolaev | ||||
| Family Hydroseraceae Nikolaev & Harwood 2002 | ||||
| Hydrosera whampoensis (A.F.Schwarz) Deby 1891 | 12, 82, 83 | M | BE | Joh (2010) |
| Order Stellarimales Nikolaev & Harwood | ||||
| Family Trigoniaceae Glezer 2019 | ||||
| Trigonium cf. curvatum A.Nair, R.Mohan, S.Shetye, S.Gazi & S.A.Jafar 2015 | 11, 79–81 | M | BE | Nair et al. (2015) |
| Class Mediophyceae Medlin & Kaczmarska 2004 | ||||
| Order Thalassiosirales Glezer & I.V.Makarova 1986 | ||||
| Family Thalassiosiraceae M.Lebour 1930 | ||||
| Thalassiosira eccentrica (Ehrenberg) Cleve 1904 | 8 | M | PL | Tomas (1997); Hoppenrath et al. (2009) |
| Order Biddulphiales Willi Krieger 1954 | ||||
| Family Biddulphiaceae Kützing 1844 | ||||
| Biddulphia biddulphiana (J.E.Smith) Boyer 1900 | 13, 84 | M/B | TP | Hoban (1983); Galland and Pennebaker (2012) |
| Order Eupodiscales V.A.Nikolaev & D.M.Harwood 2000 | ||||
| Family Odontellaceae P.A.Sims, D.M.Williams & Ashworth 2018 | ||||
| Amphitetras antediluviana Ehrenberg 1840* | 14, 85, 86 | M | BE, EL, EI | Mayama and Fujita (1995); Sims et al. (2018) |
| Odontella aurita (Lyngbye) C.Agardh 1824 | 15 | M/B | TP, EI | Sims et al. (2018); An et al. (2023) |
| Class Bacillariophyceae Haeckel 1878 | ||||
| Order Rhaphoneidales Round 1990 | ||||
| Family Psammodiscaceae Round & D.G.Mann 1990 | ||||
| Psammodiscus nitidus (Gregory) Round & Mann 1980 | 16 | M/B | TP, EI | Garcia and Talgatti (2008); Watanabe et al. (2013) |
| Order Rhabdonematales Round & R.M.Crawford 1990 | ||||
| Family Rhabdonemataceae Round & R.M.Crawford 1990 | ||||
| Rhabdonema adriaticum Kützing 1844 | 17 | M | TP, EI | Álvarez-Blanco and Blanco (2014); Witkowski et al. (2000) |
| Rhabdonema arcuatum (Lyngbye) Kützing 1844 | 18, 19 | M | TP, EI | Witkowski et al. (2000) |
| Family Grammatophoraceae Lobban & Ashworth 2014 | ||||
| Grammatophora oceanica var. subtilissima (Bailey) Grunow 1881 | 20 | M/B | TP, EI | Witkowski et al. (2000); Lee et al. (2012) |
| Grammatophora oceanica Ehrenberg 1840 | 21, 87 | M/B | EI | Álvarez-Blanco and Blanco (2014); Witkowski et al. (2000) |
| Grammatophora angulosa Ehrenberg 1840 | 22 | M/B | EI | Witkowski et al. (2000); Sato et al. (2010) |
| Grammatophora hamulifera Kützing 1844 | 23 | M/B | EI | Witkowski et al. (2000) |
| Order Cymbellales D.G.Mann 1990 | ||||
| Family Rhoicospheniaceae J.Y.Chen & H.Z.Zhu 1983 | ||||
| Rhoicosphenia abbreviata (C.Agardh) Lange-Bertalot 1980 | 24–26, 88–90 | F/B | EI | Witkowski et al. (2000); Kobayasi et al. (2006); Levkov et al. (2010); Al-Handal and Al-Shaheen (2019) |
| Order Licmophorales Round 1990 | ||||
| Family Licmophoraceae Kützing 1844 | ||||
| Podocystis spathulata (Shadbolt) Van Heurck 1896 | 27 | M | EI | Lobban et al. (2012) |
| Order Achnanthales P.C.Silva 1962 | ||||
| Family Achnanthaceae Kützing 1844 | ||||
| Achnanthes brevipes var. intermedia (Kützing) Cleve 1895 | 28–30, 91, 92 | M/B | EI | Toyoda and Williams (2004); Lee et al. (2013) |
| Achnanthes subconstricta (F.Meister) K.Toyoda 2003 | 31, 93 | F/B | EI | Toyoda et al. (2003); Lee et al. (2013) |
| Family Achnanthidiaceae D.G.Mann 1990 | ||||
| Planothidium hauckianum (Grunow) Bukhtiyarova 1999 | 32, 33, 94 | M/B | EI | Joh (2012); Kulaš et al. (2020) |
| Family Cocconeidaceae Kützing 1844 | ||||
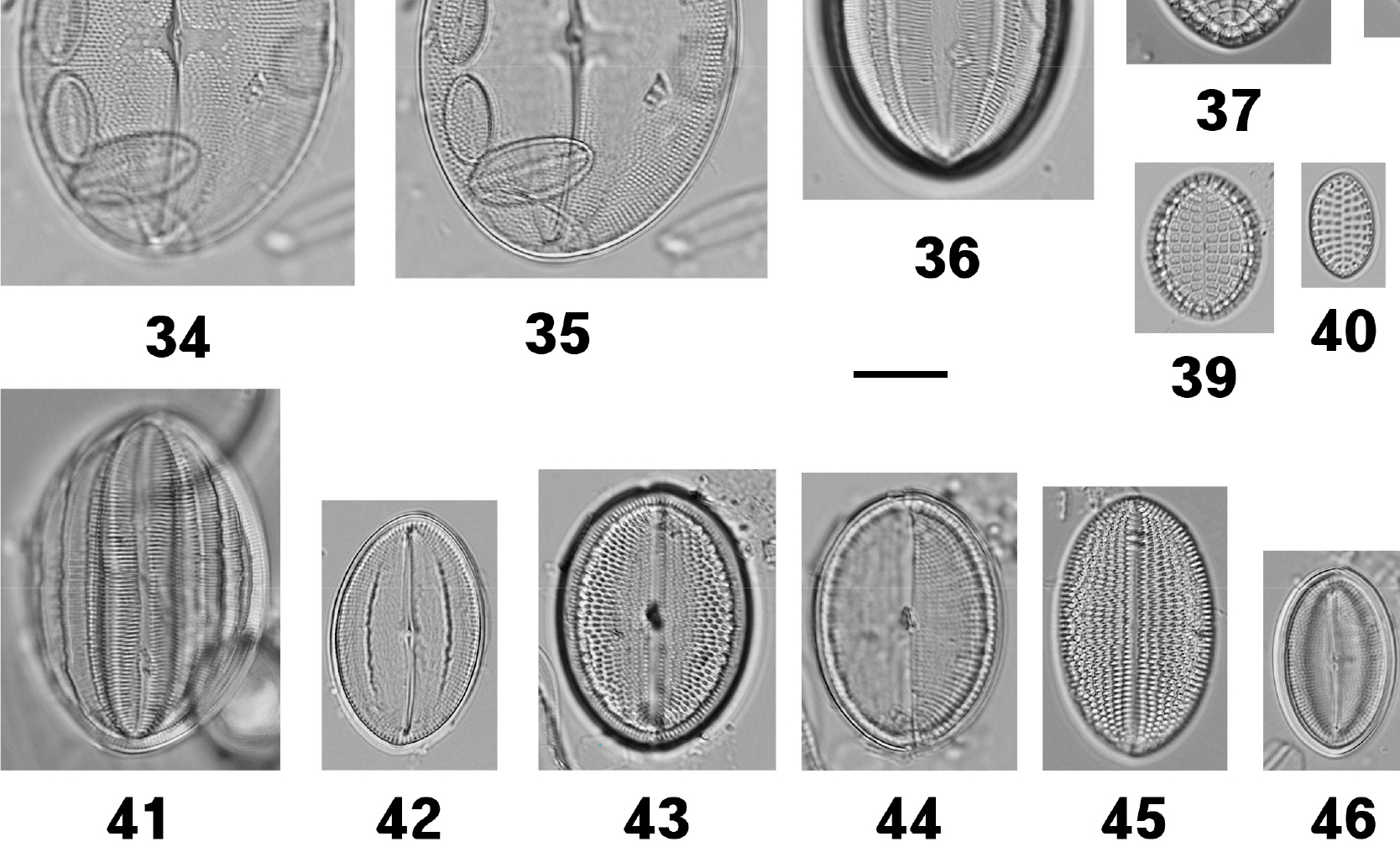
| Cocconeis pellucida Hantzsch 1863 | 34–36, 95, 96 | M | EI | Witkowski et al. (2000); Joh (2021) |
| Cocconeis scutellum Ehrenberg 1838 | 37, 97 | M/B | EI | Álvarez-Blanco and Blanco (2014); Witkowski et al. (2000); Riaux-Gobin and Romero (2003); Al-Handal and Al-Shaheen (2019); Joh (2021) |
| Cocconeis stauroneiformis H.Okuno 1957 | 38–40, 98, 99 | M/B | EI | Witkowski et al. (2000); Riaux-Gobin and Romero (2003) |
| Cocconeis krammeri Lange-Bertalot & Metzeltin | 41, 42 | M/B | EI | Álvarez-Blanco and Blanco (2014); Witkowski et al. (2000); Kim et al. (2017) |
| Cocconeis placentula Ehrenberg 1838 | 43–45 | F | EI | Al-Handal and Al-Shaheen (2019) |
| Cocconeis pseudomarginata W.Smith 1853 | 46 | M | EI | Witkowski et al. (2000) |
| Order Naviculales Bessey 1907 | ||||
| Family Amphipleuraceae Grunow 1862 | ||||
| Halamphora subholsatica (Krammer) Levkov 2009 | 47 | M/B | EP | Levkov (2009) |
| Halamphora holsatica (Hustedt) Levkov 2009 | 48, 49 | M/B | EP | Sar et al. (2003); Levkov (2009); Stepanek and Kociolek (2018) |
| Halamphora pseudoholsatica (Kobayasi & Nagumo) Levkov 2009* | 50, 100 | M/B | EP | Witkowski et al. (2000); Stepanek and Kociolek (2018) |
| Halamphora sydowii (Cholnoky) Levkov 2009* | 101 | M/B | EP | Levkov (2009); Stepanek and Kociolek (2018) |
| Amphiprora cf. pseudoduplex | 59, 105–107 | M/B | EP | Stachura-Suchoples et al. (2016); Witkowski et al. (2000); Hein et al. (2008) |
| Family Naviculaceae Kützing 1844 | ||||
| Navicula flagellifera Hustedt 1939 | 51, 52 | M/B | EP | Witkowski et al. (2000); Hein et al. (2008) |
| Navicula perminuta Grunow 1880 | 53, 102 | F/B | EP | Fukushima et al. (1992); Kang et al. (1999); Witkowski et al. (2000) |
| Seminavis robusta D.B.Danielidis & D.G.Mann 2002 | 54 | M/B | EP | Wachnicka and Gaiser (2007); Hein et al. (2008); Al-Handal and Al-Shaheen (2019) |
| Trachyneis velata (A.W.F.Schmidt) Cleve 1894 | 55 | M | EP | Witkowski et al. (2000); Siqueiros Beltrones et al. (2016) Lobban and Tharngan (2025) |
| Family Diadesmidaceae D.G.Mann 1990 | ||||
| Luticola sp. | 56, 103 | M/B | EP | |
| Luticola mutica (Kützing) D.G.Mann 1990 | 57, 58, 104 | F/S | EP | Álvarez-Blanco and Blanco (2014); Al-Handal and Al-Shaheen (2019) |
| Family Pleurosigmataceae Mereschowsky 1903 | ||||
| Pleurosigma elongatum W.Smith 1852 | 60, 61 | M/B | EP | Taylor et al. (2007); Al-Handal and Al-Shaheen (2019) |
| Order Bacillariales Hendey 1937 | ||||
| Family Bacillariaceae Ehrenberg 1831 | ||||
| Nitzschia incurva Grunow 1878 | 62 | M/B | TP, EP | Al-Yamani and Saburova (2019); Lobban and Witkowski (2023) |
| Nitzschia gracilis Hantzsch 1860 | 63 | F/B | EP | Siver et al. (2005); Kihara et al. (2009); Hamsher et al. (2016) |
| Nitzschia dissipata (Kützing) Grunow 1862 | 64, 65 | F/B | EP | Watanabe (2005); Tuji (2016); Al-Handal and Al-Shaheen (2019) |
| Nitzschia inconspicua Grunow 1862 | 66, 67, 108 | F/B | EP | Watanabe (2005); Trobajo et al. (2013); Al-Handal and Al-Shaheen (2019) |
| Tryblionella coarctata f. densestriata (Hustedt) A.M.M.Freitas & M.P.Lobo 2018* | 68, 109 | M/B | EP | Álvarez-Blanco and Blanco (2014) |
| Order Surirellales D.G.Mann 1990 | ||||
| Family Entomoneidaceae Reimer 1975 | ||||
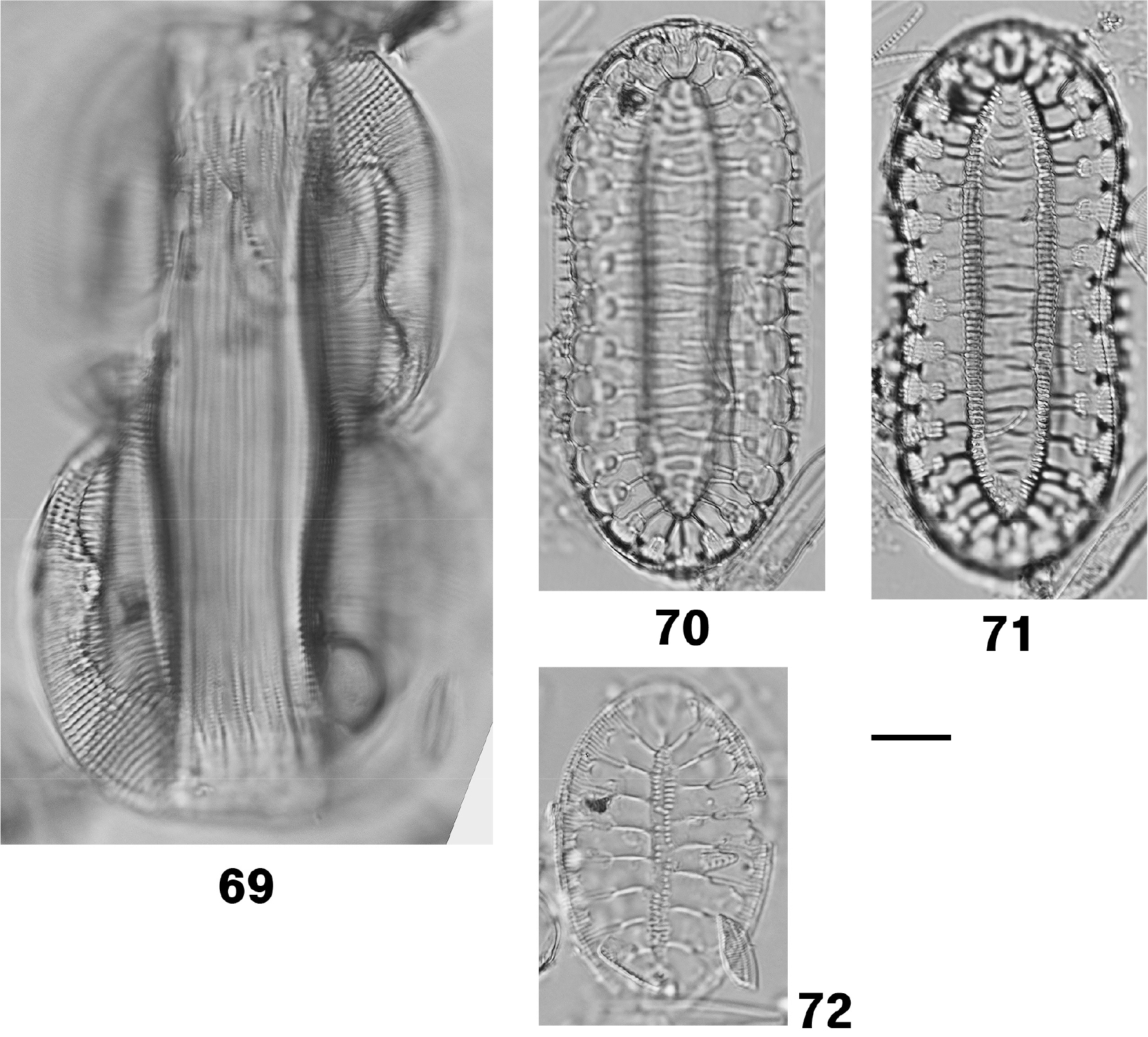
| Entomoneis japonica (Cleve) K.Osada 2002* | 69, 110, 111 | M/B | EP | Osada and Kobayasi (1985); Kobayasi et al. (2006) |
| Family Surirellaceae Kützing 1844 | ||||
| Surirella taeniata Hustedt ex Simonsen 1987 | 70, 71 | F/B | EP | Schmidt (1925); Park et al. (2017) |
| Surirella recedens A.W.F.Schmidt 1875 | 72 | F/B | EP | Park et al. (2012); Park et al. (2021) |
Trigonium cf. curvatum A.Nair, R.Mohan, S.Shetye, S.Gazi & S.A.Jafar 2015
Key reference: Nair et al. (2015), Figs. 1-12.

Description: Frustules robust, tripolar. Valves triangular, with distinctly concave sides and apices raised into prominent pseudocelli. Valve face ornamented with polygonal, projecting loculate areolae, 2–3 µm in diameter; each loculus bordered by 4–6 internal puncta. Areolae towards the apices progressively reduced, forming hyaline pseudocellular fields. Central valve region bearing a cluster of approximately 12 short, tubular rimoportulae. Valve mantle vertical, with areolate pattern continuous from the valve face (Figs. 11 and 79–81).


Figs. 17–33.
Light micrographs of eleven diatom species. Fig. 17. Rhabdonema adriaticum. Figs. 18 and 19. R. arcuatum. Fig. 20. Grammatophora oceanica var. subtilissima. Fig. 21. G. oceanica. Fig. 22. G. angulosa. Fig. 23. G. hamulifera. Figs. 24–26. Rhoicosphenia abbreviata. Fig. 27. Podocystis spathulata. Figs. 28–30. Achnanthes brevipes var. intermedia. Fig. 31. A. subconstricta. Figs. 32 and 33. Planothidium hauckianum. Scale bar is 10 µm

Figs. 47–68.
Light micrographs of sixteen diatom species. Fig. 47. Halamphora subholsatica. Figs. 48 and 49. H. holsatica. Fig. 50. H. pseudoholsatica. Figs. 51 and 52. Navicula flagellifera. Fig. 53. N. perminuta. Fig. 54. Seminavis robusta. Fig. 55. Trachyneis velata. Fig. 56. Luticola sp. Figs. 57 and 58. L. mutica. Fig. 59. Amphiprora pseudoduplex. Figs. 60 and 61. Pleurosigma elongatum. Fig. 62. Nitzschia incurva. Fig. 63. N. gracilis. Figs. 64 and 65. N. dissipata. Figs. 66 and 67. N. inconspicua. Fig. 68. Tryblionella coarctata f. densestriata. Scale bar is 10 µm

Figs. 73–84.
Scanning electron micrographs of seven diatom species. Figs. 73 and 74. Melosira inflexa. Fig. 75. M. nummuloides. Fig. 76. Paralia guyana. Figs. 77 and 78. Arachnoidiscus ehrenbergii. Figs. 79–81. Trigonium cf. curvatum. Figs. 82 and 83. Hydrosera whampoensis. Fig. 84. Biddulphia biddulphiana. Scale bars: 20 µm (76–79, 82–84); 10 µm (73–75, 81); 5 µm (80)
Remarks: The present material is provisionally assigned to Trigonium cf. curvatum, originally described from Prydz Bay, Antarctica (Nair et al. 2015), and later revised to its current orthography in accordance with Articles 23.5 and 62.2 of the International Code of Nomenclature (ICN). The specimens examined in this study closely match the original description regarding valve outline, the structure of the loculate areolae, and the presence of a central cluster of rimoportulae. Nair et al. (2015) differentiated T. curvatum from T. arcticum partly based on the absence of central labiate processes in the latter species. However, subsequent research has revealed that T. arcticum can exhibit numerous labiate processes (e.g. 25–80 per valve) (Hoban 1983; Ferrario and Ferreyra 1984; Gibson and Mahoney 1984), suggesting that this characteristic is variable and thus unreliable for taxonomic differentiation. Moreover, taxonomic delimitation within biddulphioid diatoms, including the genus Trigonium, has historically been challenging, as closely related taxa are often confused or misidentified due to overlapping morphological traits (Gibson and Mahoney 1984). These findings highlight the limitations of relying on single or variable morphological characters for species-level identification within this group.
Despite the inherent complexities, the present material can be differentiated from T. arcticum based on a consistent set of morphological characteristics. Specifically, the valves exhibit distinctly concave sides, feature projecting loculate areolae measuring less than 5 µm in diameter with 4–6 internal puncta, and possess relatively narrow girdle bands. Conversely, T. arcticum is characterized by straight to convex valve margins, flattened areolae exceeding 5 µm in diameter with a greater number of puncta, and broader girdle bands. The specimens analyzed in this study correspond closely to this morphotype in all primary diagnostic traits.
Nevertheless, certain differences were observed, including a smaller valve size (apical axis approximately 99.8 µm compared to 175–360 µm in Antarctic populations) and a reduced number of labiate processes (approximately 12 versus 20–24). This variation may reflect size reduction during vegetative reproduction or regional morphological variability, both of which have been documented in biddulphioid diatoms.
T. curvatum is presently considered taxonomically unresolved in AlgaeBase. Despite the general morphological agreement observed, and recognizing the inherent complexity of this group, the specimens examined herein are provisionally assigned to Trigonium cf. curvatum. Definitive clarification of its taxonomic status necessitates comprehensive investigations, including detailed analyses of girdle ultrastructure and molecular markers such as 18S rRNA and rbcL genes.
Distribution and ecology: Nair et al. (2015) described Trigonium curvatum as a large epiphytic–benthic diatom predominantly found in marginal-marine environments, especially within sediments associated with sea ice in Antarctic regions. Although initially reported from polar habitats, species within the T. arcticum complex have since been identified across a wide range of geographic locations, encompassing temperate and tropical environments (Krayesky et al. 2009; Gupta and Das 2020). This evidence suggests that their distribution is not exclusively limited to cold or polar conditions.
From an ecological perspective, the presence of such taxa beyond their originally documented range should be approached with caution. The distribution of benthic and epiphytic diatoms is heavily influenced by local environmental factors, substrate properties, and biological interactions, often leading to assemblages that are highly dynamic and context-dependent (Hinode et al. 2021). Furthermore, species such as T. arcticum have been observed in a diverse array of marine habitats, including epiphytic environments (Tiffany and Lange 2002), indicating a broader ecological tolerance than previously recognized.
In the present study, the sampling site is affected by submarine groundwater discharge (SGD), resulting in physicochemical conditions that differ markedly from those of the adjacent coastal waters. The recorded temperature (16.6°C) and salinity (2 psu) reflect a pronounced freshwater influence, leading to the establishment of a relatively stable microhabitat. These localized conditions may promote the persistence of taxa commonly associated with cooler or marginal-marine environments. Nevertheless, additional ecological and physiological data are required to comprehensively assess this interpretation.

Figs. 85–96.
Scanning electron micrographs of seven diatom species. Figs. 85 and 86. Amphitetras antediluviana. Fig. 87. Grammatophora oceanica. Figs. 88–90. Rhoicosphenia abbreviata. Figs. 91 and 92. Achnanthes brevipes var. intermedia. Fig. 93. A. subconstricta. Fig. 94. Planothidium hauckianum. Figs. 95 and 96. Cocconeis pellucida. Scale bars: 20 μm (85); 10 μm (87–89, 91–93, 95); 5 μm (86, 90, 94, 96)
Amphitetras antediluviana Ehrenberg 1840
Key references: Mayama and Fujita (1995), Figs. 1-11; Sims et al. (2018), Figs. 95–100.
Synonyms: Triceratium antediluvianum (Ehrenberg) Grunow 1868; Biddulphia antediluviana (Ehrenberg) Van Heurck 1885; Odontella antediluviana (Ehrenberg) M.Peragallo 1903.
Description: Frustules cuboidal in girdle view. Valves square to slightly rhomboidal, with straight to slightly concave margins and broadly rounded to weakly produced angles; valve width (side length) 58.8 µm. Central annulus circular to oval, from which robust costae radiate towards the corners, occasionally bifurcating distally. Costae interconnected by vimines, forming large quadrangular areolae, 1.5–3 µm in diameter, spaced c. 4 µm apart. Valve face separated from mantle by a distinct marginal rim, terminating in a peripheral flange. Each corner bearing an ocellus composed of a silica ring enclosing densely packed porelli, externally ornamented with granules. Areolae internally occluded by two types of cribra: (i) a poroid mesh supported by delicate, bifurcating struts, and (ii) a cribrum formed by thick, spathulate plates producing T-shaped slits. Girdle bands numerous, open, each with a uniseriate row of rectangular areolae. Raphe absent.
Remarks: Amphitetras antediluviana was initially described as a fossil species by Ehrenberg (1840) and has subsequently been extensively characterized through both light and scanning electron microscopy (Mayama and Fujita 1995; Jahn and Kusber 2006; Sims et al. 2018; Nevrova et al. 2024). The specimens examined in the present study correspond closely with these previous descriptions in terms of overall valve morphology, ocellus structure, and internal areolar architecture.
This species can be differentiated from morphologically similar biddulphioid diatoms, including those of the genus Biddulphia, by a consistent set of diagnostic characteristics. Valves of A. antediluviana are typically quadrate, featuring four well-developed ocelli at the corners and a central annulus from which costae radiate, creating a regular quadrangular areolation pattern (Jahn and Kusber 2006; Sims et al. 2018). In contrast, species within the genus Biddulphia generally display more rounded valve outlines, lack distinct ocelli, and exhibit differences in the arrangement of costae and girdle structures (Sims et al. 2018). Additional distinguishing characteristics include the internal occlusion of areolae by cribra and the absence of a raphe system, both of which align with previous descriptions (Mayama and Fujita 1995; Sims et al. 2018). These combined features offer a reliable basis for differentiating A. antediluviana from superficially similar taxa.
Distribution and ecology: Amphitetras antediluviana was initially described by Ehrenberg (1840) as a marine species and is presently recognized as an extant taxon exhibiting a broad, nearly cosmopolitan distribution. This species has been documented in diverse coastal and marine environments throughout Europe, North and South America, Africa, Asia, and Australasia, reported under its accepted name as well as various synonyms, including Triceratium antediluvianum and Biddulphia antediluviana.
In Asia, A. antediluviana has been recorded in the Northwest Pacific region, encompassing Chinese coastal waters such as the Bohai Sea and adjacent northern coastal areas (Liu 2008), the Sea of Japan along the Russian Far Eastern coast, including the Peter the Great Bay region (Gail 1950), and Japanese coastal environments such as Arikawa Bay in Nagasaki Prefecture (Hinode et al. 2021). These observations suggest that the species is well established in the temperate marginal seas of East Asia.
The current documentation from Korean coastal waters expands the recognized distribution of A. antediluviana within East Asia. The SGD environment at Chunggulmul, characterized by the interaction between groundwater and seawater, likely offers favorable microhabitats for marine and euryhaline diatom taxa, including this species.
Halamphora pseudoholsatica (T.Nagumo & H.Kobayasi) J.G.Štěpánek & Kociolek 2018
Key references: Witkowski et al. (2000), p. 148, pl. 161, Figs. 5 and 6; Stepanek and Kociolek (2018), p. 33–34, pl. 31, Figs. 1-4.
Basionym: Amphora pseudoholsatica T.Nagumo & H.Kobayasi
Description: Valves strongly dorsiventral, semi-elliptical to semi-lanceolate, 15.7–24.8 µm long, 3.7–6.9 µm wide. Dorsal margin smoothly arched; ventral margin straight to slightly convex; apices narrowly sub-capitate, deflected ventrally. Junction between valve face and dorsal mantle marked by a distinct marginal silica ridge; dorsal mantle perpendicular. Mantle areolae arranged differently from those on the valve face. Dorsal striae coarse, distinctly punctate, uniseriate, 12 in 10 µm; several central dorsal striae shortened or fused, forming a small siliceous nodule. Ventral striae very fine, uniseriate, 17 in 10 µm. Internally, marginal ridge absent; dorsal virgae coalescing into a small hyaline fascia beneath the central nodule (Figs. 50 and 100).
Remarks: Published populations of Halamphora pseudoholsatica generally exhibit valve lengths ranging from 23 to 53 µm and breadths between 4.5 and 8 µm (Witkowski et al. 2000; Stepanek and Kociolek 2018). However, smaller valves measuring 18–21 µm in length and 3.3–4.5 µm in breadth have been documented in the Black Sea (Zidarova et al. 2022). The specimens examined in the present study (15.7–24.8 µm × 3.7–6.9 µm) extend the known lower size range of the species and likely correspond to late-series valves formed during size diminution.
The specimens can be reliably identified by a consistent set of diagnostic characteristics, which include a well-developed dorsal marginal ridge, coarse punctate uniseriate dorsal striae, and extremely fine ventral striae. These features are stable and have been highlighted in previous electron microscopy studies as essential criteria for the identification of H. pseudoholsatica (Witkowski et al. 2000; Stepanek and Kociolek 2018).
Halamphora pseudoholsatica has historically been misidentified with several related taxa. It can be distinguished from H. holsatica by its narrower valve width (typically less than 7 µm), the presence of a distinct dorsal marginal ridge, and finer ventral striae. In contrast, H. holsatica lacks the dorsal ridge and exhibits coarser ventral striae, measuring 16–18 in 10 µm (Sar et al. 2003). H. holsaticoides is characterized by a thin silica strip adjacent to the dorsal raphe and a more perpendicular mantle. Meanwhile, H. subholsatica lacks the dorsal ridge and displays ventral striae that are interrupted at the mid-valve; some authors consider this taxon conspecific with H. acutiuscula, which is distinguished by a more convex dorsal margin and more radiate dorsal striae (Sar et al. 2003).
Distribution and ecology: Halamphora pseudoholsatica was initially described from intertidal mudflats in Tokyo Bay and is currently recognized in temperate to subtropical coastal regions, including the Baltic Sea, North Sea, North Atlantic estuaries, and the Black Sea. This species has been characterized as a marine to brackish benthic diatom inhabiting coastal environments (Witkowski et al. 2000), exhibiting a broad salinity tolerance ranging from approximately 10 to 35 psu (Stepanek and Kociolek 2018).
The species is typically observed as an epipelic or epipsammic taxon on muddy or sandy substrates and has also been documented as epiphytic on macroalgae. This study constitutes the first recorded occurrence of H. pseudoholsatica in Korea.
Halamphora sydowii (Cholnoky) Levkov 2009
Key references: Levkov (2009), p. 232–234, pl. 105, Figs. 7–10, pl. 242, Figs. 1–4>; Stepanek and Kociolek (2018), p. 77, pl. 71, Figs. 5–8, pl. 74, Figs. 1–6.
Basionym: Amphora sydowii Cholnoky
Description: Valves semi-lanceolate, distinctly dorsiventral, 28 µm long, 4.7 µm wide. Dorsal margin strongly convex; ventral margin straight to slightly concave; apices short-protracted, rostrate to sub-capitate, deflected ventrally. Raphe positioned on a narrow, slightly elevated dorsal ledge. Proximal raphe ends closely appressed, expanded into small central pores, bent dorsally before slightly diverging; distal ends curved dorsally into the apices. Dorsal striae radiate, 17 in 10 µm, biseriate near the raphe ledge, becoming uniseriate towards the dorsal margin. Ventral striae very fine, uniseriate, 26 in 10 µm, each composed of a single elongate areola (Fig. 101).

Figs. 97–111.
Scanning electron micrographs of eleven diatom species. Fig. 97. Cocconeis scutellum. Figs. 98 and 99. C. stauroneiformis. Fig. 100. Halamphora pseudoholsatica. Fig. 101. H. sydowii. Fig. 102. Navicula perminuta. Fig. 103. Luticola sp. Fig. 104. L. mutica. Figs. 105–107. Amphiprora pseudoduplex. Fig. 108. Nitzschia inconspicua. Fig. 109. Tryblionella coarctata f. densestriata. Figs. 110 and 111. Entomoneis japonica. Scale bars: 20 µm (110); 10 µm (99–101, 105, 106, 111); 5 µm (97, 98, 102–104, 107–109)
Remarks: Halamphora sydowii is most likely to be confused with other small, strongly dorsiventral species; however, it can be distinguished by a consistent combination of valve breadth, stria structure, and raphe morphology.
This species differs from H. coffeaeformis in possessing dorsal striae that become uniseriate toward the margin and ventral striae composed of a single elongate areola, whereas H. coffeaeformis maintains biseriate dorsal striae and ventral striae consisting of 2–4 poroids (Levkov 2009). Additionally, Halamphora hybrida is narrower (3–4 µm in width), displays uniformly uniseriate dorsal striae, and has nearly straight proximal raphe ends, in contrast to H. sydowii, which is broader and exhibits proximal fissures distinctly deflected toward the dorsal side (Levkov 2009). Halamphora holsatica exhibits biseriate dorsal ornamentation and frequently biseriate striae, features that are absent in H. sydowii (Stepanek and Kociolek 2018). In contrast, Halamphora witkowskii is characterized by coarser dorsal striae (12–15 in 10 µm), consistently biseriate striae, denser ventral striae that may be partially biseriate, and a broader valve width ranging from 6 to 8 µm (Van de Vijver et al. 2014). Halamphora acutiuscula is narrower, measuring 3–4 µm, possesses long-capitate apices, nearly straight proximal raphe ends, and lacks the longitudinal rib found in H. sydowii (Yılmaz et al. 2025). Lastly, Halamphora veneta is distinguished by a more pronounced concave ventral margin, markedly down-curved apices, and finer dorsal striae (18–24 in 10 µm) (Levkov 2009).
The specimen analyzed in this study corresponds closely to the diagnostic characteristics of H. sydowii; however, the identification is based on a single valve.
Distribution and ecology: Halamphora sydowii has been documented in a diverse array of freshwater to brackish environments throughout Europe and Asia, encompassing alkaline rivers, canals, reservoirs, and spring systems (Levkov 2009; Van de Vijver et al. 2014). This species demonstrates a wide ecological tolerance and has been observed in conditions ranging from oligohaline to mesohaline (Yılmaz et al. 2025). The species has been documented in a variety of hydrological environments, including mineral-rich thermal waters, spring-fed streams, and mildly brackish coastal springs, demonstrating significant ecological adaptability to low-salinity conditions. This study constitutes the first recorded occurrence of H. sydowii in Korea.
Luticola sp.
Description: Valve rectangular in girdle view. Valves linear- to elliptic-lanceolate with broadly rounded, non-protracted apices, 21 µm long, 7.1 µm wide. Axial area narrow, straight, slightly widening towards a panduriform central area, bordered on each side by one to two enlarged areolae. Striae radiate throughout, 22 in 10 µm; each stria composed of three to four round to slightly elongate areolae, continuing onto the mantle as a single row with corresponding number of pores. A slit-like stigma present submarginally on the secondary side, accompanied externally by a minute occluded marginal pore. On the primary side, two to three shallow pit-like depressions present midway between the sternum and valve margin. Raphe filiform; proximal raphe ends attenuated, weakly deflected towards the primary side and then slightly recurved towards the stigma, lying nearly parallel to the apical axis. Distal raphe fissures hooked towards the stigma, terminating on the valve face (Figs. 56 and 103).
Remarks: The current taxon can be assigned to the genus Luticola due to the presence of a single isolated stigma, radiate striae composed of distinct areolae, and a raphe system exhibiting differentiated proximal and distal endings, which align with the contemporary circumscription of the genus (Da Silva-Lehmkuhl et al. 2019; Lokhande et al. 2021).
Morphologically, the specimens exhibit overall similarity to Luticola hunanensis, particularly in valve outline characterized by broadly rounded, non-protracted apices, the presence of a slit-like stigma associated with a marginal pore, and the general configuration of the central area. However, a notable discrepancy is evident in the morphology of the proximal raphe endings. According to the protologue of L. hunanensis, the proximal raphe fissures are distinctly hook-shaped, displaying an approximately right-angled deflection toward the stigma-bearing side (Liu et al. 2017). In contrast, the specimens under study possess proximal endings that are attenuated and follow a gently S-shaped course. Given that proximal raphe morphology is considered a critical diagnostic character within Luticola, this difference precludes a definitive assignment to that species.
In addition to L. hunanensis, the specimens exhibit general morphological characteristics consistent with several small Luticola species, including L. mutica, L. frequentissima, and L. goeppertiana. The present material differs from L. mutica by possessing a distinctly panduriform central area bordered by enlarged areolae and a higher stria density (22 in 10 µm). L. frequentissima differs in the configuration of the central area and the morphology of the distal raphe endings, which are typically more extended and oriented differently (Noga et al. 2017). In contrast, the distal fissures in the present material hook toward the stigma and terminate on the valve face. Although L. goeppertiana exhibits a comparable general outline, it differs in central area development and proximal raphe morphology (Pavlov et al. 2009). Additional distinctions are apparent when compared with L. beyensii, particularly regarding central area structure and raphe configuration (Van de Vijver et al. 2002).
Despite these comparisons, the observed combination of characteristics—including a panduriform central area, relatively high stria density, a slit-like stigma accompanied by a marginal pore, and attenuated proximal raphe endings—does not correspond exactly to any previously described species. This challenge aligns with recent research indicating that the genus Luticola is morphologically complex and taxonomically challenging, exhibiting high species diversity and frequently overlapping diagnostic features, especially in freshwater to low-salinity habitats (Da Silva-Lehmkuhl et al. 2019; Bagmet et al. 2023).
From an ecological standpoint, the presence of this taxon within a coastal SGD system necessitates careful interpretation. Luticolahunanensis was initially described from freshwater environments (Liu et al. 2017); however, the current specimens were obtained under conditions of strong freshwater influence (approximately 2 psu), suggesting that the site represents a localized low-salinity microhabitat. Previous research has demonstrated that Luticola species inhabit a broad spectrum of salinity regimes, including marine and brackish waters, indicating a wider ecological tolerance than previously recognized (Rybak et al. 2021). Furthermore, the genus has been documented across diverse hydrological contexts and biogeographic regions, reflecting substantial ecological plasticity and dispersal capacity (Lokhande et al. 2021).
In this context, the observed population may represent (i) a resident population adapted to low-salinity conditions within the SGD system, (ii) allochthonous input originating from groundwater or inland freshwater sources, or (iii) a distinct yet morphologically similar taxon inhabiting transitional habitats.
Due to the discrepancy observed in a critical diagnostic character and the limited availability of specimens, the material is provisionally classified as Luticola sp. Comprehensive further studies, encompassing the collection of additional specimens, detailed scanning electron microscopy analyses of multiple valves, and molecular data acquisition, are required to clarify its taxonomic status.
Tryblionella coarctata f. densestriata Álvarez-Blanco & S.Blanco 2014
Reference: Álvarez-Blanco and Blanco (2014), pl. 40: Figs. 7–10, pl. 87: Fig. 3
Description: Valves linear-panduriform, distinctly constricted at mid-length and widening towards broadly cuneate, short-rostrate apices; a longitudinal fold present along the valve face. Valve 20.1 µm long, 7.7 µm wide. Raphe keel strongly eccentric, with a distinct central nodule. Fibulae 15 in 10 µm. Striae punctate, 23 in 10 µm, arranged in transverse rows and interrupted by the longitudinal fold. Areolae coarse, arranged in distinct tiers; puncta appearing enlarged where crossing the fold (Figs. 68 and 109).
Remarks: Tryblionella coarctata f. densestriata exhibits the panduriform valve outline, a strongly eccentric raphe keel, and a prominent longitudinal fold, consistent with the nominate taxon. However, it is distinguished by its smaller valve dimensions (10–23 µm × 5–7 µm compared to 25–140 µm × 11–22 µm) and a higher density of transapical striae (22–30 per 10 µm versus ≤ 18). Additionally, the fibulae number ranges from 13 to 16 per 10 µm.
Among related taxa, Psammodictyon panduriforme var. continuum also exhibits panduriform valves and punctate striae; however, it is distinguished by the presence of a hyaline half-valve, coarser striation, and a less pronounced longitudinal fold (Round et al. 1990). In contrast, Nitzschia panduriformis and N. constricta display a similar valve outline but possess a central (rather than strongly eccentric) raphe keel and lack the distinct longitudinal fold that interrupts the striae (Snoeijs and Balashova 1998).
Distribution and ecology: Tryblionella coarctata f. densestriata was initially described as a free-living epilithic species inhabiting coastal rocks near El Mojón, Murcia, Spain (Álvarez-Blanco and Blanco 2014). Subsequently, it has also been documented on stones in the brackish waters of Ereğli Bay along the Turkish Black Sea coast (Dirican et al. 2022). The specimens obtained from the oligohaline coastal spring Chunggulmul on Jeju Island constitute the first recorded occurrence of this taxon in Korea and the broader northwestern Pacific region. In addition to its free-living occurrences, this taxon has been documented as an intracellular symbiont of the foraminifer Pararotalia calcariformata along the Israeli coast (Schmidt et al. 2018). These observations suggest that the species inhabits a variety of coastal and brackish environments and may occupy both benthic substrates and symbiotic niches.
Entomoneis japonica (Cleve) K.Osada 2002
Key references: Osada and Kobayasi (1985), p. 215–224, pl. 1–5: Figs. 1–30; Kobayasi et al. (2006), p. 144, pl. 179, Figs. 1–8.
Basionym: Amphiprora alata var. japonica Cleve 1894.
Synonym: Entomoneis alata var. japonica (Cleve) Osada & Kobayasi 1985.
Description: Frustules panduriform, slightly twisted along the apical axis; in girdle view violin-shaped and distinctly constricted at mid-length. Valves linear-lanceolate with acutely pointed apices, 103.4 µm long. Striae parallel, biseriate, 11 in 10 µm. Each stria composed of two rows of poroid areolae, externally occluded by dome-shaped hymenes perforated by scattered elongate pores. Raphe canal forming a strongly S-shaped, wing-like keel elevated above the valve surface. The junction between the keel and valve body defined by a row of basal fibulae, appearing distinctly bisinuous on either side of the central nodule. Cingulum composed of five girdle bands; areolae occluded by hymenes with circular pores centrally and elongate pores towards the margins (Figs. 69, 110 and 111).
Remarks: This species is distinguished from related taxa by a combination of morphological features, including a violin-shaped valve outline, a consistently bisinuous junction line, and relatively coarse, biseriate striae (11–12 in 10 µm). These characters differentiate it from E. paludosa, which has finer, uniseriate striae (21–26 in 10 µm), and from E. punctulata and E. aequabilis, which exhibit much higher stria densities (≥ 32 in 10 µm) (Mejdandžić et al. 2017).
In addition, the scattered arrangement of hymen pores on the valve face contrasts with the centrally aligned pattern observed in E. alata (Kobayasi et al. 2006). Observations from clonal cultures indicate that valve length may be reduced to approximately 55 µm without affecting stria density, keel morphology, or junction-line structure, supporting the stability of these diagnostic characters (Osada and Kobayasi 1985).
Distribution and ecology: The species is common in brackish-water habitats across Japan (Kobayasi et al. 2006). Field collections from tidal-flat muds and estuarine sediments indicate a benthic, attached life form. Clonal cultures demonstrate growth at salinities of 15–25 psu, indicating an affinity for moderate brackish conditions (Osada and Kobayasi 1985). Accordingly, Entomoneis japonica is regarded as a euryhaline, epipelic to epilithic species typical of estuarine environments. This study represents the first record of this species in Korea.
4. Conclusion
This study documents the diatom flora of the Chunggulmul submarine groundwater discharge (SGD) spring on Jeju Island and provides the first floristic account of diatoms from this unique habitat in Korea. A total of 48 taxa belonging to 28 genera were identified based on LM and SEM observations. Among them, five species are reported for the first time in Korea, while two taxa (Trigonium cf. curvatum and Luticola sp.) remain unresolved at the species level. The assemblage included taxa with freshwater, brackish, and marine affinities, reflecting the variable environmental conditions associated with groundwater–seawater mixing in the spring. Because studies on diatom communities from SGD environments in Korea remain limited, the present work provides baseline floristic information for this habitat. Further investigations across multiple seasons and sites would help to better understand the ecological dynamics and biodiversity of diatoms in groundwater-influenced coastal systems.
