

1. 서 론
2. 연구방법
연구지역
측정 및 분석
3. 결 과
이화학적 특성
부착조류의 현존량 및 출현특성
기수갈고둥의 출현밀도 및 분포 특성
기수갈고둥의 밀도와 환경요인과의 관계
4. 고 찰
1. 서 론
기수갈고둥(Clithon retropictum)은 연체동물문(Mollusca) 복족강(Gastropoda) 고리갈고둥목(Cycloneritida) 갈고둥과(Neritidae)에 속하는 종으로, 일본, 중국 남부, 대만 등의 동아시아 지역에 분포한다(Noseworthy et al. 2013). 우리나라에서는 제주연안, 남해안 및 동해 남부연안의 기수역에서 관찰되며, 약한 유속과 자갈 등으로 구성되어 부유물질 농도가 상대적으로 적은 하천의 기수역에 서식한다(오와 김 2018). 기수갈고둥이 서식하고 있는 기수역은 하천의 정비공사 및 매립 등으로 서식처가 파괴됨에 따라 자연개체군의 감소가 예상되어 환경부에서는 멸종위기야생생물 2급으로, 해양수산부에서는 해양보호생물로 지정하여 관리하고 있다. 그럼에도 불구하고 기수갈고둥은 상대적으로 개체 크기가 작고 돌 아래에 부착하여 서식하기 때문에 쉽게 눈에 띠지 않으며 인지도가 낮을 뿐만 아니라 지속적인 하천 정비공사 등으로 자연개체군 유지에 위협이 되고 있다.
동아시아에만 분포하는 기수갈고둥의 특성으로 인해 일본에서 많은 연구가 수행되었다. 일본의 주요 하천에서 분포, 이동 및 서식생태 등에 관한 연구(Kumazawa et al. 1991; Nishiwaki et al. 1991a; Hirata et al. 1992; Furujo and Tomiyama 2000; Ohara and Tomiyama 2000; Shigemiya and Kato 2001; Kobayashi and Iwasaki 2002; Hirata et al. 2014; Miyajima and Wada 2014; Haraguchi and Tomiyama 2018)가 수행되었고, 기타 산란특성(Nishiwaki et al. 1991b)과 미토콘드리아 DNA (Fukumori et al. 2016) 연구가 있다. 반면 우리나라에서는 초등학교 교원이 전국과학전람회에서 기수갈고둥의 서식지 환경 및 생태에 관한 발표(박과 이 2008; 조와 황 2010)를 시작으로 2010년대부터 관심을 갖기 시작하면서 각종 언론보도를 통해 서식처가 발견되고 있다. 본격적인 연구는 제주 연안에서 기수갈고둥의 형태 및 출현(Noseworthy et al. 2012; Noseworthy et al. 2013), 전사체(transcriptome) 및 미토콘드리아 DNA 분석(Park et al. 2016; Cho et al. 2017), 서식처의 보호(Choi et al. 2018) 및 환경영향 평가기법(오와 김 2018)에 관한 연구와 경상남도 하천에서 기수갈고둥의 분포 현황(홍 등 2019)에 관한 보고가 있다. 특히 염분 변화가 극심한 하천에서 기수갈고둥의 생태적 특성에 관한 연구로는 경남 거제시 연초천에서 염분과 토양입도 등의 환경조건에 따른 크기와 밀도의 영향 분석(이 등 2018)으로, 주요 활동시기인 5월부터 11월까지 4회에 걸쳐 연구한 결과 뿐이다.
최근 멸종위기 야생생물 및 해양보호생물에 대한 관심이 높아지고 있으나, 상대적으로 크기가 작으면서도 개발 및 접근이 용이한 지역(조간대 및 하천)에 서식하는 고둥류나 게류는 위협 요인이 상존하고 있어 더 많은 관심과 연구가 필요하다. 특히, 바다와 연결된 하천의 기수역에 서식하는 기수갈고둥의 자연개체군 확대를 위해서는 서식지 보존과 더불어 서식처의 이화학적 특성과 먹이가 되는 생물에 대한 전반적인 이해를 파악하는 등 실내 인공증식을 위한 기초자료의 확보가 시급하다. 본 연구에서는 남해안에서 서식 개체수가 많은 2개 하천을 대상으로 이화학적 환경 인자와 먹이생물인 부착조류의 출현 특성을 분석하여 개체군의 밀도와 치패의 가입에 영향을 주는 인자를 분석하고자 하였다.
2. 연구방법
연구지역
기수갈고둥 서식처의 이화학적 환경특성과 서식밀도를 분석하기 위해 국내 주요 서식처인 봉강천과 미룡천을 선정하였다. 봉강천은 전라남도 보성군 회천면에 위치하는 지방 하천으로, 길이는 2.4 km, 유역 면적은 6.10 km2이고, 미룡천은 경상남도 고성군 삼산면에 위치하며 하천 길이는 2 km, 유역 면적은 3.67 km2이다(Fig. 1).
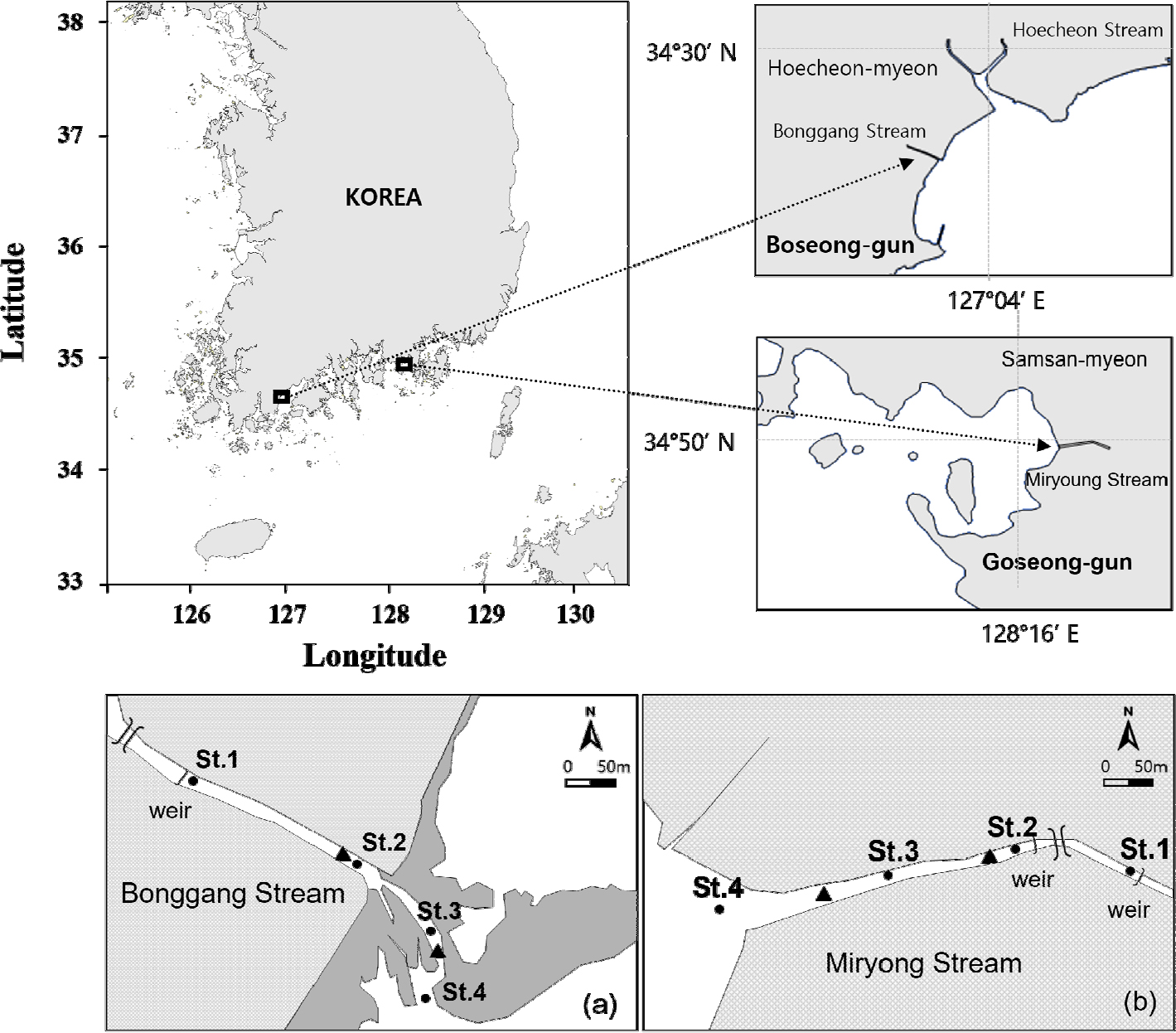
연구대상 2개 하천을 대상으로 기수갈고둥의 서식이 확인된 가장 위쪽 정점과 가장 아래쪽 정점 사이에 2개의 정점을 추가하여 총 4개 정점을 선정하였다. 봉강천은 하천 종단부에서 내측으로 약 160 m 지점에 인공구조물인 보를 사이에 두고 기수갈고둥의 서식 여부가 확연히 구분되었고(St. 1), 하천 종단부 기저에 하천 폭이 넓어지는 지점(St. 2), 하천 폭이 좁아지면서 유속이 급격히 빨라지는 지점(St. 3), 하천 종단부 외측 지점(St. 4)으로 구분하였다(Fig. 1a). 미룡천은 하천 종단부에서 내측으로 200 m 부근과 300 m 부근에 보가 있으나, 300 m 부근 보를 기준으로 기수갈고둥의 서식 여부가 구분되었고(St. 1), 두 번째 보를 지나 유속이 빨라지는 지점(St. 2), 하천 폭이 넓어지는 지점(St. 3), 하천 종단부 외측(St. 4)으로 구분하였다(Fig. 1b). 또한 조석에 따른 수온, 염분, 수위의 일간변화를 분석하기 위해 봉강천은 수로가 넓어지는 내측 지점과 유속이 빨라지는 외측 지역으로 구분하였고(Fig. 1a), 미룡천은 유속이 빨라지는 내측 지점과 수로가 넓어지는 외측 지점으로 구분하였다(Fig. 1b).
측정 및 분석
본 연구는 해당 지방환경청으로부터 사전 허가를 통해 진행되었으며, 봉강천은 2018년 3월부터 2019년 2월까지, 미룡천은 2019년 3월부터 2020년 2월까지 매월 1회 현장측정과 서식밀도를 분석하였다. 각각의 정점에서 다항목수질측정기(YSI Proplus, USA)를 이용하여 수온 및 염분과 수소이온농도 및 용존산소량을 현장에서 측정하였다. 또한 각 하천의 2개 정점을 추가로 선정하여 조석에 따른 수온, 염분, 수위 변화를 측정하였는데, 연중 조석간만의 차가 큰 백중사리 기간(8월) 중 하루를 정해 10분 간격으로 24시간 동안 실시간 측정센서(Onset Hobo, USA)를 이용하여 측정하였다. 기준이 되는 조위 변화자료는 국립해양조사원에서 발표한 것으로, 봉강천은 전라남도 녹동항, 미룡천은 경상남도 남포항의 조석표를 참고하였다.
기수갈고둥의 주요 먹이를 분석하기 위해 각 정점에서 기질의 표면으로부터 부착조류의 시료를 채집하였다. 기질의 선택은 물 흐름과 일치하고, 조석에 관계없이 항시 잠겨있는 평평한 10 cm 이상의 자연석을 선택하였으며, 채집용 솔로 기질 표면의 25 cm2를 긁고 증류수로 씻어 정량 채집하였다. 채집된 시료 중 종 동정을 위한 시료는 채집 즉시 Lugol’s 용액으로 고정한 후 연구실로 운반하여 광학현미경(Axioskop, Zeiss) 200~400배 하에서 정량 분석하였다. Chlorophyll-a는 동정용 시료와 동일한 방법으로 채집한 후 현장에서 필터한 GF/C 여과지를 냉장 상태로 운반한 다음 아세톤추출법으로 색소를 추출하고 형광광도계(10-AU, Turner Designs, USA)로 측정하였다.
기수갈고둥의 정량채집 및 측정을 위해 각각의 정점에서 50 cm × 50 cm 방형구 내에 존재하는 모든 개체를 1회 채집한 후 개체별로 크기(각고 및 각폭, 0.01 mm)와 무게(0.001 g)를 측정하였다. 측정이 끝난 개체는 본래의 서식 위치에 방사하였다.
수온 및 염분 등의 수질환경과 부착조류의 분류군별 현존량이 기수갈고둥의 개체군 밀도와 크기에 미치는 영향을 분석하기 위해 통계 프로그램(SPSS v.24)를 이용하여 상관관계 분석을 하였고, 정점별 크기 분포의 유의성을 분석하기 위해 One-way ANOVA 분석을 실시하였다.
3. 결 과
이화학적 특성
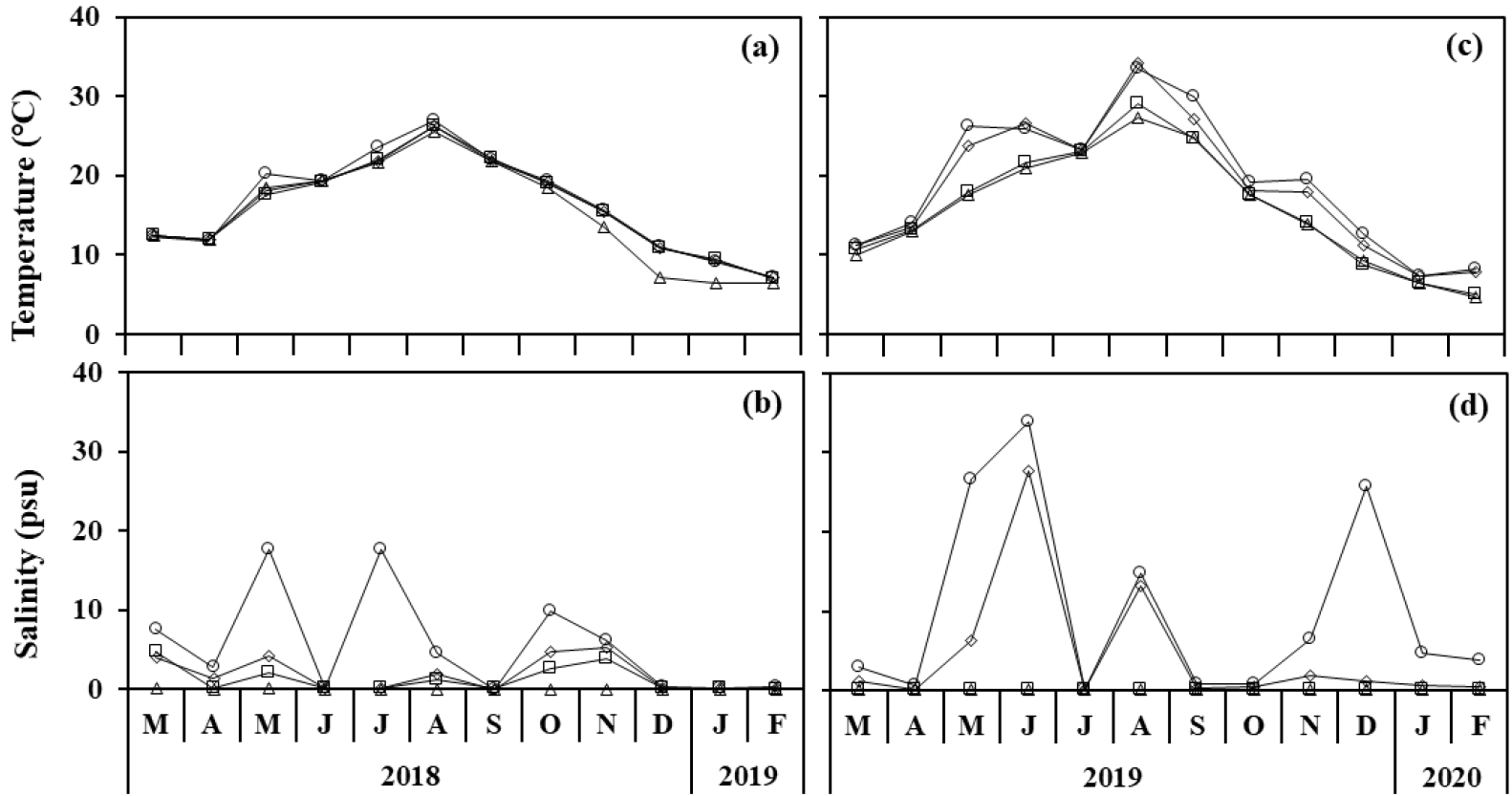
봉강천에서 수온 변화는 월 평균 6.9~26.2°C로, 수위가 낮고 담수 환경인 최 내측 정점 1이 다른 정점에 비해 변화 폭이 크게 나타났다(Fig. 2a). 염분은 측정 당일 조석의 차이에 따라 변화를 보였는데, 시기에 따라 평균 0.1~6.0 psu이었고, 최대 17.7 psu로 가장 바깥쪽 정점에서 7월에 나타났다(Fig. 2b). 미룡천의 수온과 염분 변화는 계절 및 정점에 따라 봉강천과 유사한 경향을 보였고, 월 평균 수온은 6.5~31.0°C로 봉강천에 비해 변화 폭이 더 컸다(Fig. 2c). 염분 역시 측정 당일의 조석에 따라 다르게 나타났는데, 월 평균 0.1~15.5 psu의 범위를 보였고, 각 정점에서 최대 염분은 최 외측의 정점 4에서 33.9 psu (6월)이었다(Fig. 2d).
8월 백중사리 기간 중 24시간 동안 10분 간격으로 측정된 실시간 수온, 염분, 수위 변화자료는 Fig. 3과 같다. 봉강천의 수온 변화는 내측 측정지점에서 23.8~29.0°C, 외측 측정지점에서 23.7~28.6°C의 범위로 두 측정지점의 차이는 크지 않았으며, 고조시에 수온이 증가하였다(Fig. 3a). 염분은 내측 측정지점에서 0.4~23.4 psu, 외측 측정지점에서 0.8~27.7 psu의 범위로 하천 종단부에 가까운 외측 측정지점에서 높은 염분이 측정되었다(Fig. 3b). 수온과 달리 외측 측정지점에서는 창조와 낙조 사이에 0.9~ 27.7 psu의 급격한 염분 변화를 보인 반면, 내측 지점에서는 저저조와 고고조 사이 창조시에만 0.4~23.4 psu의 급격한 승강 변화가 관측되었고, 고고조 이후 낙조와 창조에서는 최저 8.5 psu로 그 변화가 적었다. 수위 변화는 고고조와 저고조에 따라 다른 양상을 보였으며, 외측 측정지점에서 최대 1.2m로 수위 변화 폭이 컸다(Fig. 3c).
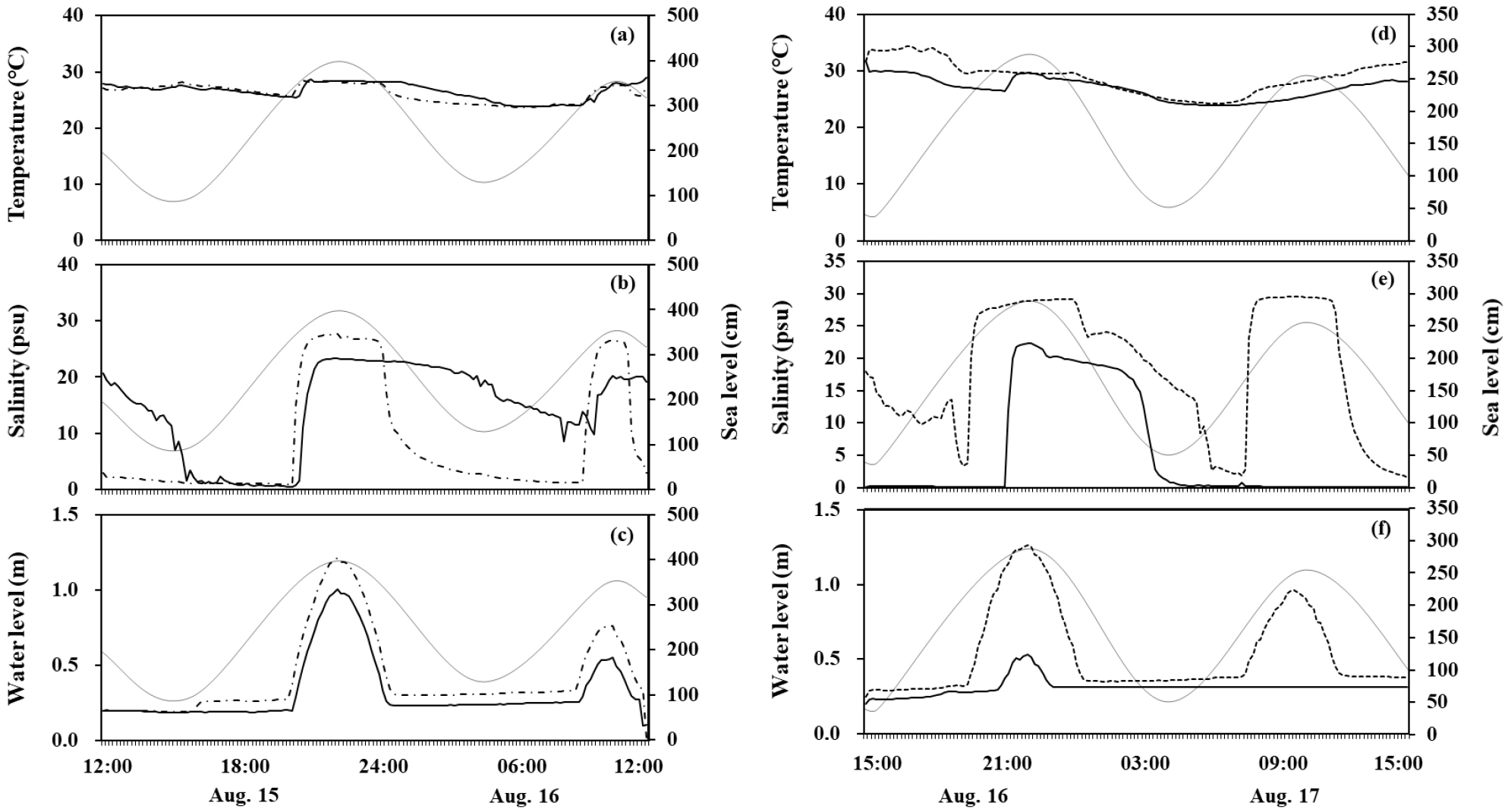
Fig. 3.
Time series of temperature, salinity, and water level fluctuation during 24 hours in Bonggang Stream (a–c) and Miryong Stream (d–f). Dotted line, bold line, and thin line indicated outer point, inner point, and tide level, respectively. Data for tidal levels observed Nockdong Port (a–c), Cheollanam-do and Nampo Port (d–f), Kyeongsangnam-do, respectively
미룡천의 수온은 내측 측정지점에서 23.9~31.7°C, 외측 측정지점에서 24.2~34.4°C의 범위로 외측 지점에서 주간에 해당하는 시간대에 다소 높게 관측되었다(Fig. 3d). 염분은 내측 측정지점에서 0.1~22.3 psu, 외측 측정지점에서 1.7~29.6 psu의 범위로 하천 종단부에 가까운 외측 측정지점에서 높은 염분을 나타냈다(Fig. 3e). 내측 측정지점에서는 고고조의 창조와 낙조에서만 염분변화가 관측되었고, 저고조에서는 변화가 관측되지 않았다. 외측 측정지점에서는 고고조 이후 낙조에서 염분이 서서히 낮아진데 반해, 저고조 이후 낙조에서는 급격히 염분이 낮아졌다. 수위는 내측 지점에서는 고고조에서만 0.5 m까지 수위가 증가하였고, 외측 지점에서는 고고조에서 1.3 m, 저고조에서 1.0 m로 수위가 증가하였다(Fig. 3f).
부착조류의 현존량 및 출현특성
봉강천에서 연간 출현한 부착조류는 총 6개 분류군 45종이었으며, 정점 1에서 35종으로 많은 출현종수를 보였고, 모든 정점에서 규조류가 주요 우점 분류군이었다(Table 1). 평균 현존량은 정점 4에서 가장 낮았고, 정점 1과 4는 남조류가, 정점 2와 3은 규조류의 현존량이 많았다. 미룡천에서는 연간 5개 분류군 51종이 출현하였으며, 출현종수는 정점별 차이가 크기 않았다. 정점별 평균 현존량은 정점 4에서 가장 높았고, 정점 1을 제외한 모든 정점에서 규조류가 가장 높은 밀도로 출현하였다.
Table 1.
The number of species (N) and mean abundance (A, x103 cells cm-2) of epilithic algae from monthly sampling sites in Bonggang Stream and Miryong Stream
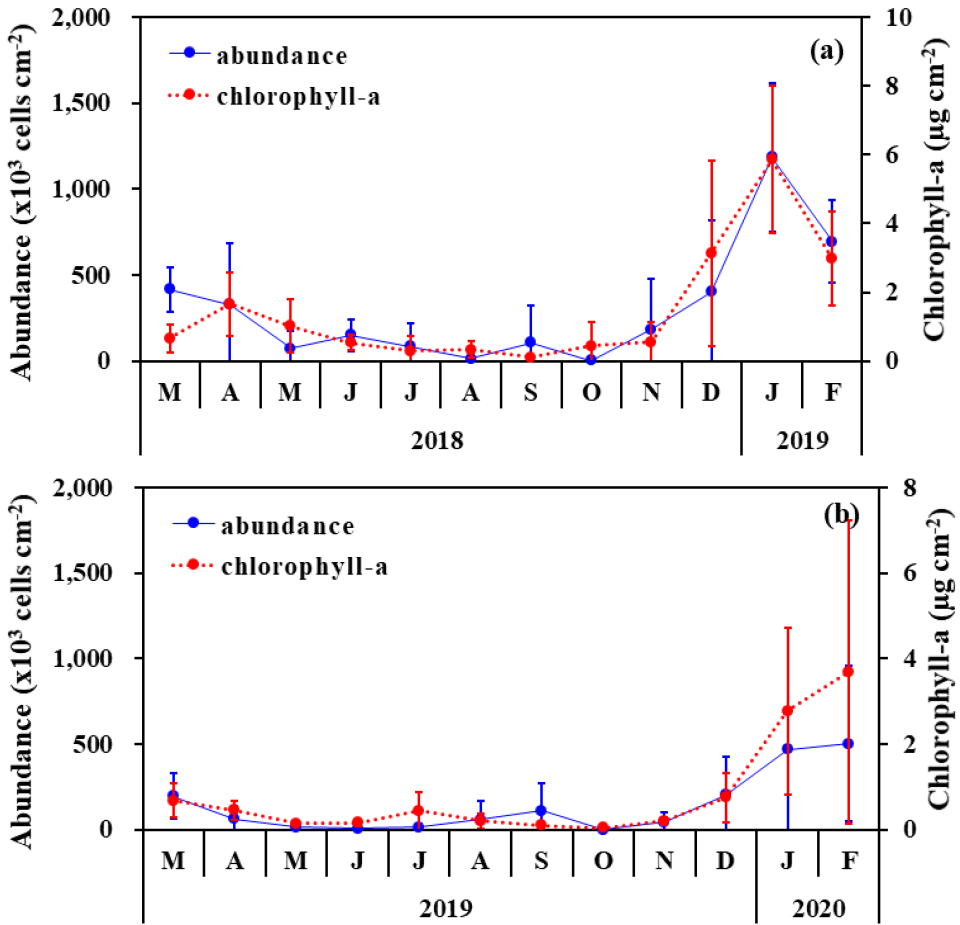
월별 부착조류의 평균 현존량과 엽록소 a를 비교하였다(Fig. 4). 월별 평균 현존량은 봉강천에서 3.8~1184.7 × 103 cells cm-2의 범위로 2018년 10월에 가장 낮고, 2019년 1월에 가장 높았다. 미룡천에서는 0.9~503.5 × 103 cells cm-2의 범위로 2019년 10월에 가장 낮고, 2020년 2월에 가장 높았다. 엽록소 a의 월별 평균 농도는 봉강천에서 0.12~ 5.86 µg cm-2의 범위로 2018년 9월에 가장 낮고 2019년 1월에 가장 높았으며, 미룡천에서는 0.05~3.69 µg cm-2의 범위로 2019년 10월에 가장 낮고 2020년 2월에 가장 높았다.
두 지역 모두 규조류, 남조류, 녹조류가 계절에 따라 우점을 달리하였는데, 봉강천은 7월에서 11월, 미룡천에서는 6월에서 11월까지 남조류 및 녹조류가 우점하였고, 그 외 시기에는 규조류가 우점하였다(Table 2). 두 지역의 정점별 부착조류 우점종을 과(Family) 수준으로 구분한 결과(Fig. 5), 봉강천의 최 내․외측인 정점 1과 4에서 남조류인 Oscillatopiaceae과(우점종은 Phormidium spp., Lyngbya sp., Phormidium tenue)의 부착조류가 각각 58.2%, 49.0%로 우점하였고, 중간 지역의 정점 2와 정점 3에서는 규조류의 Naviculaceae과(우점종은 Navicula gregaria, Naviula sp.)가 각각 30.3%와 28.8%로 가장 우점하는 분류군이었다(Fig. 5b1–b4). 미룡천은 정점 1과 정점 2에서 남조류인 Oscillatopiaceae과(우점종은 Phormidium spp., Lyngbya sp., P. tenue, Oscillatoria sp.)의 점유율이 각각 85.1%, 33.8%로 우점하였고, 정점 3과 정점 4에서는 Bacillariaceae과(우점종은 N. inconspicua, Nitzschia sp.)가 우점하였는데, 각각 42.2%와 47.2%를 차지하였다(Fig. 5m1–m4).
Table 2.
Relative abundance (%) of epilithic algal groups in Bonggang Stream and Miryong Stream (BA: Bacillariophyceae, CY: Cyanophyceae, CH: Chlorophyceae)
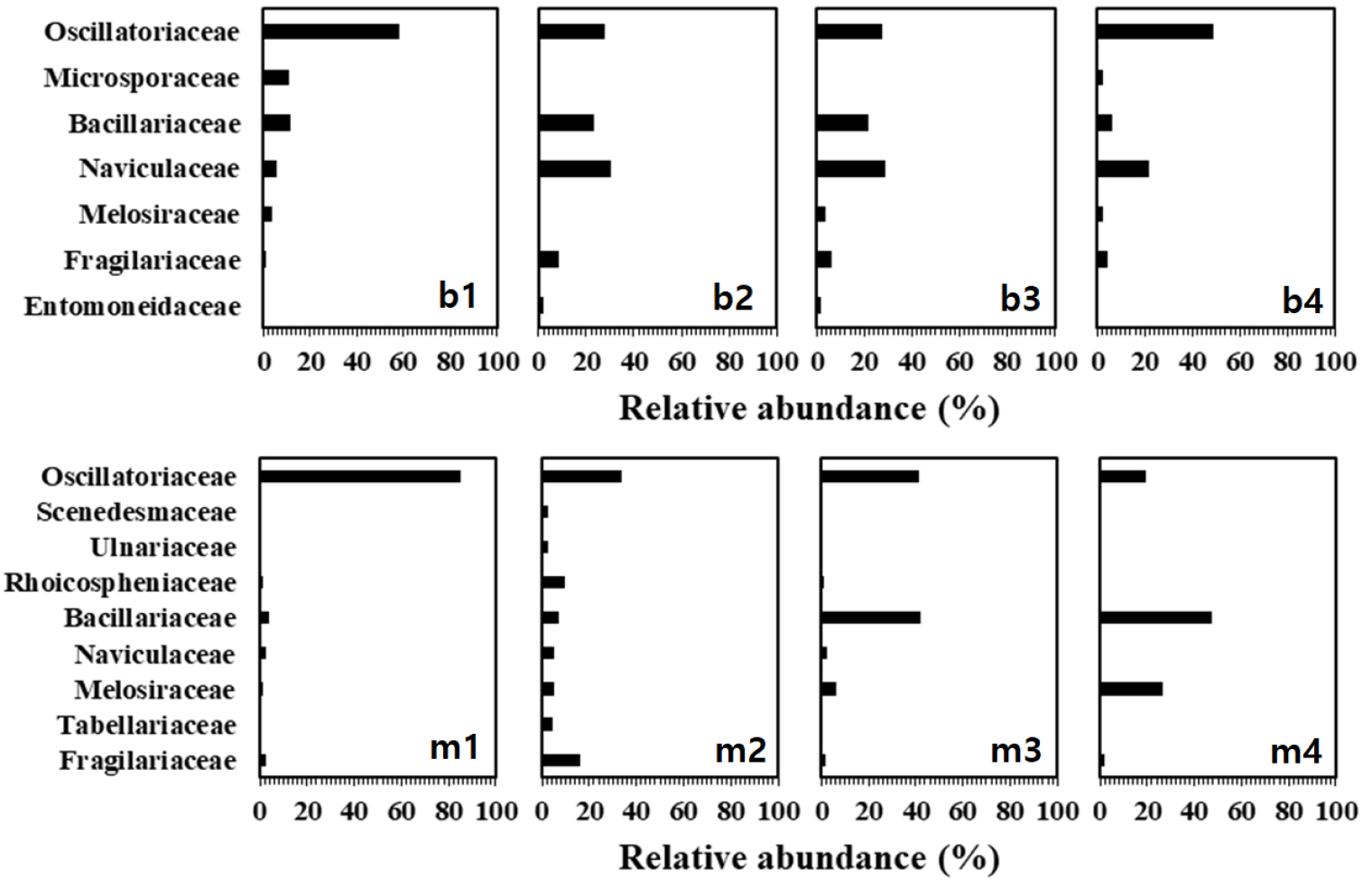
기수갈고둥의 출현밀도 및 분포 특성
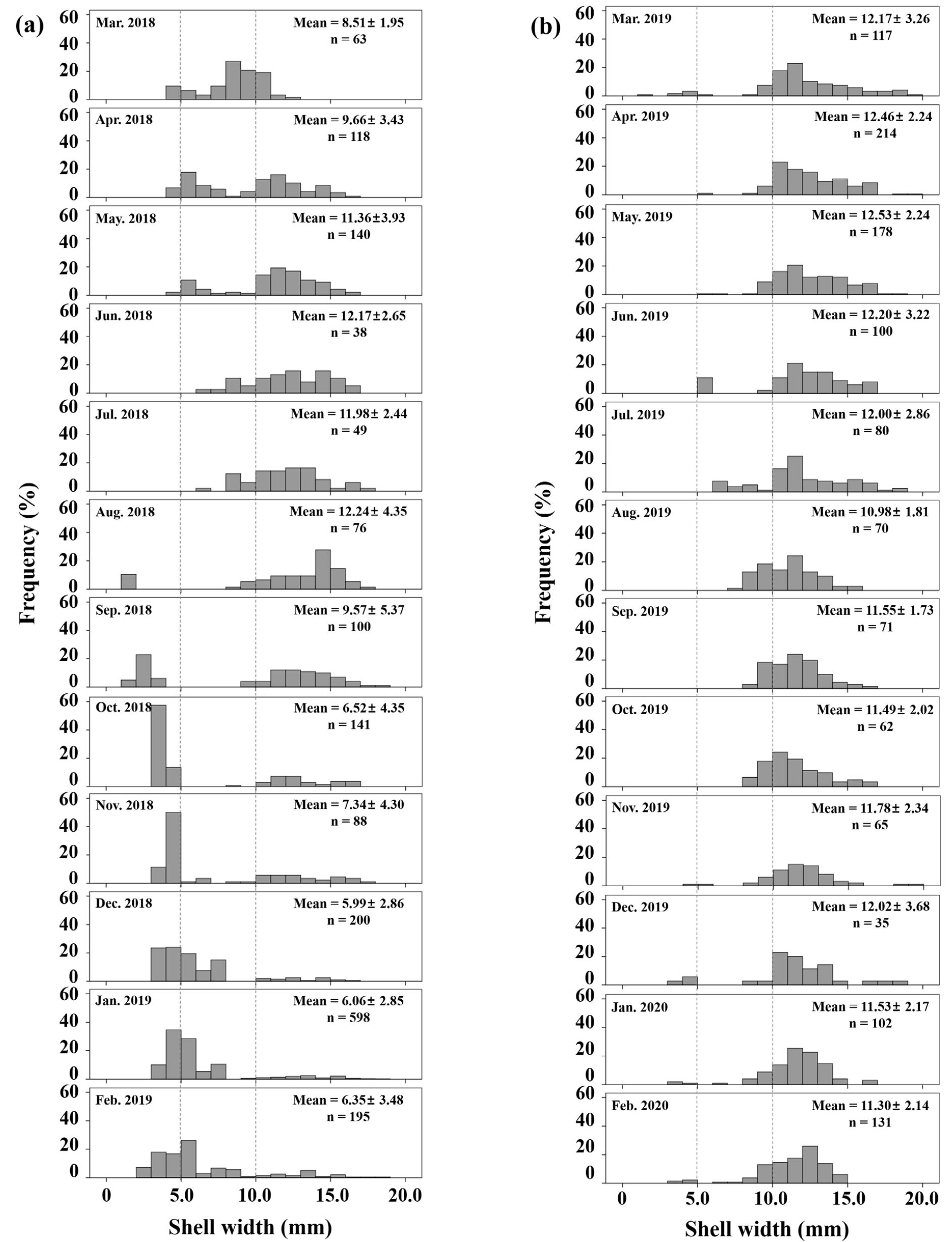
봉강천에서 기수갈고둥의 연간 평균 출현밀도는 정점 1에서 139.3 inds. m-2, 정점 2에서 360.7 inds. m-2, 정점 3에서 361.0 inds. m-2, 정점 4에서 347.8 inds. m-2로, 평균 302.2 inds. m-2가 출현하였다(Fig. 6a). 시기별로는 8월 이후 작은 개체의 가입으로 급격히 증가하였고, 최대 출현밀도는 2,288 inds. m-2로 1월에 정점 3에서 나타났다. 평균 각폭과 전중량의 월 변화를 보면(Fig. 6b and c), 전체적으로 8월 이후에 감소하였고 정점에 따라 변화 폭이 크게 나타났다. 미룡천의 연간 평균 출현밀도는 정점 1에서 7.0 inds. m-2, 정점 2에서 75.7 inds. m-2, 정점 3에서 328.7 inds. m-2, 정점 4에서 676.0 inds. m-2로, 평균 271.8 inds. m-2이었다(Fig. 6d). 2019년 5월과 2020년 1~2월에 높은 밀도를 보였으며, 정점 1에서 출현밀도가 적었으나 각폭과 전중량은 높게 나타났다(Fig. 6e and f).
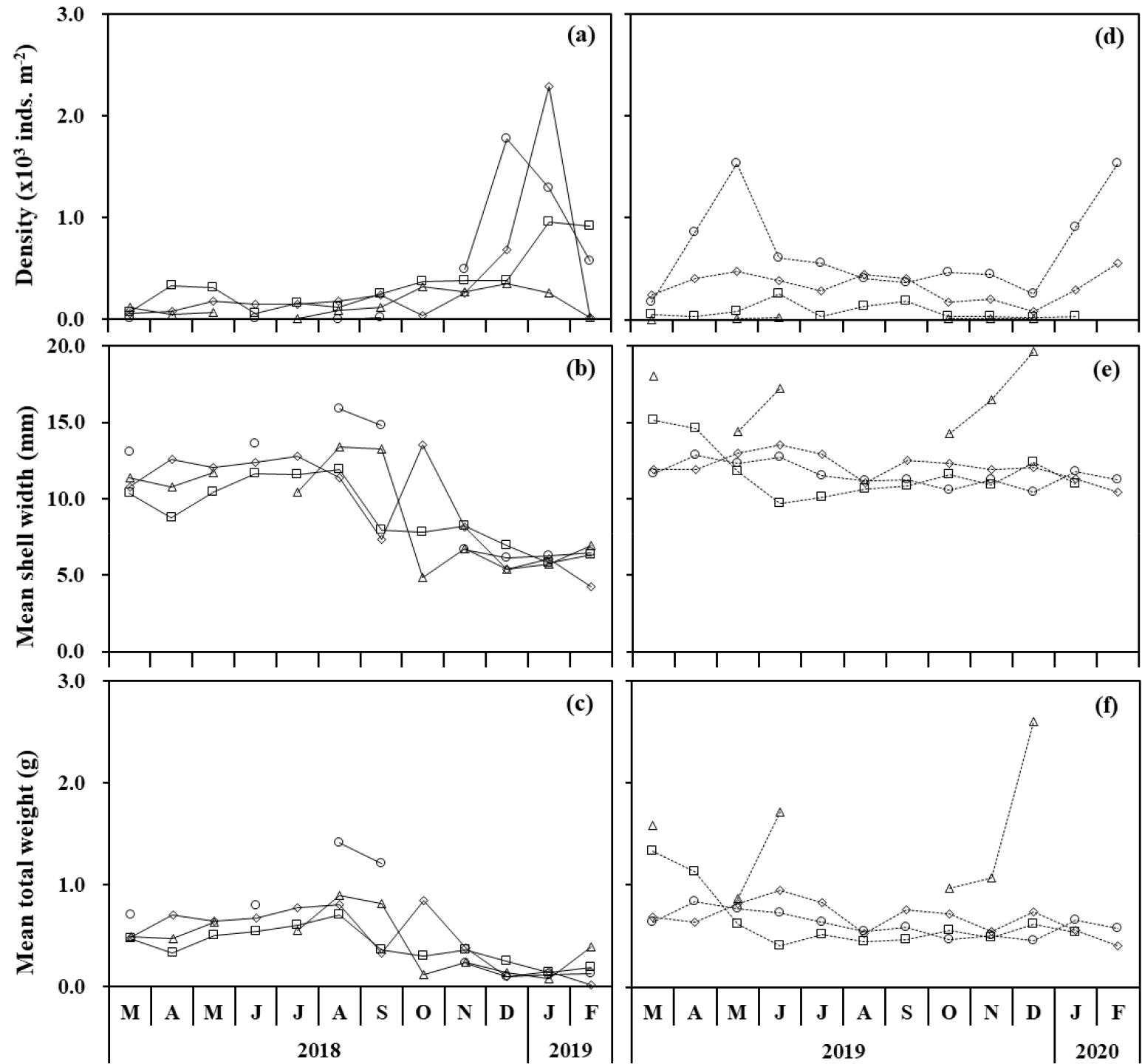
정점에 따른 월별 평균 각폭과 전중량 변화를 보면(Fig. 6), 봉강천의 경우 가장 바깥쪽 정점 4에서는 3월부터 9월까지 출현밀도가 낮아 평균 크기 및 무게가 높게 나타났으나, 11월부터 2월까지는 높은 출현밀도에 의해 평균 크기 및 무게가 급격히 감소하였다(Fig. 6a–c). 미룡천의 경우, 가장 안쪽의 정점 1에서 출현밀도가 매우 낮았고 평균 크기와 무게는 다른 정점에 비해 매우 높게 나타났다(Fig. 6d–f). 각 하천에서 정점별로 출현한 모든 개체를 대상으로 각폭, 각고 및 전중량에 대해 정점별 평균 간의 차이를 분석하였다(Table 3). 봉강천에서 평균 각폭은 6.50±2.79 mm (정점 4)~8.26±4.47 mm (정점 1)의 범위로 가장 바깥 쪽의 정점 4에서 평균 크기 및 무게가 작고, 다른 정점과 유의적인 차이를 보였다(p < 0.05). 반면 미룡천의 경우, 평균 각폭은 11.35±3.33 mm (정점 2)~16.50±2.91 mm (정점 1)의 범위로 평균 11.96±2.49 mm로 나타나 봉강천에 비해 더 컸으며, 봉강천의 출현양상과 같이 가장 안쪽 정점에서 개체 크기가 유의적으로 크게 나타났다(p < 0.05).
Table 3.
Shell width, shell height, and total weight of Clithon retropictum at sampling sites in Bonggang Stream and Miryong Stream
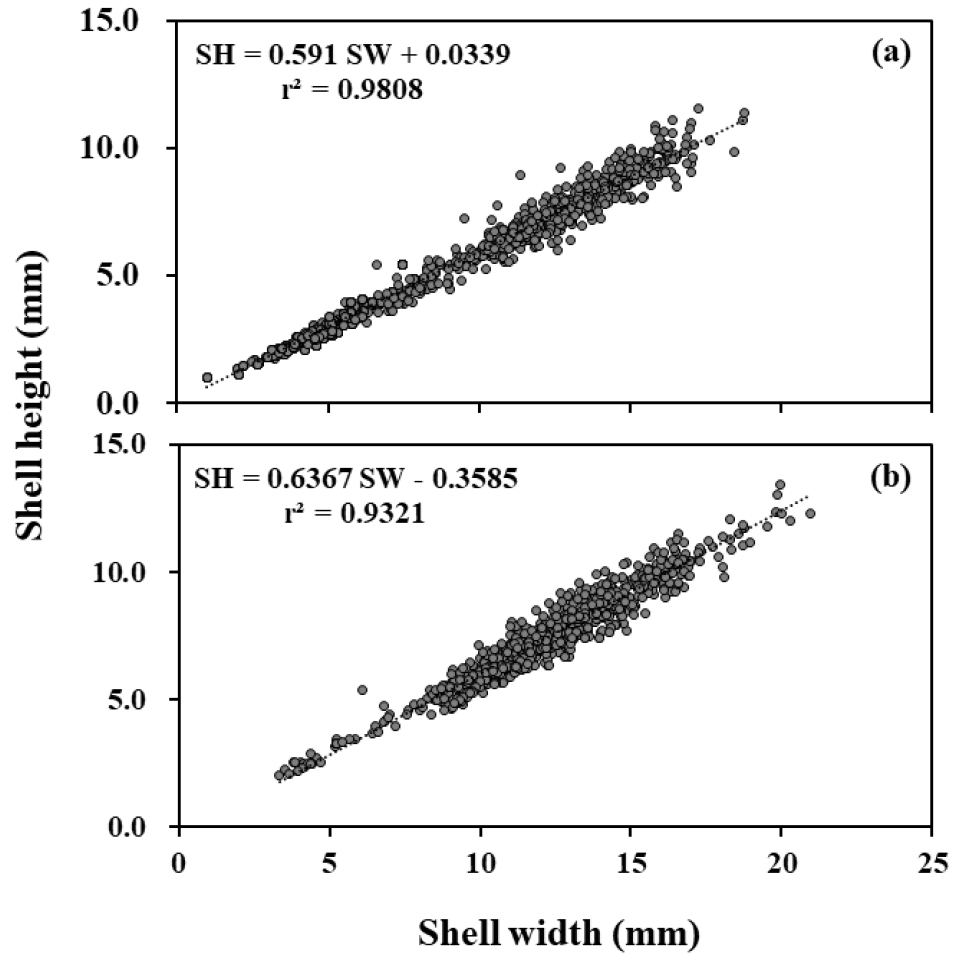
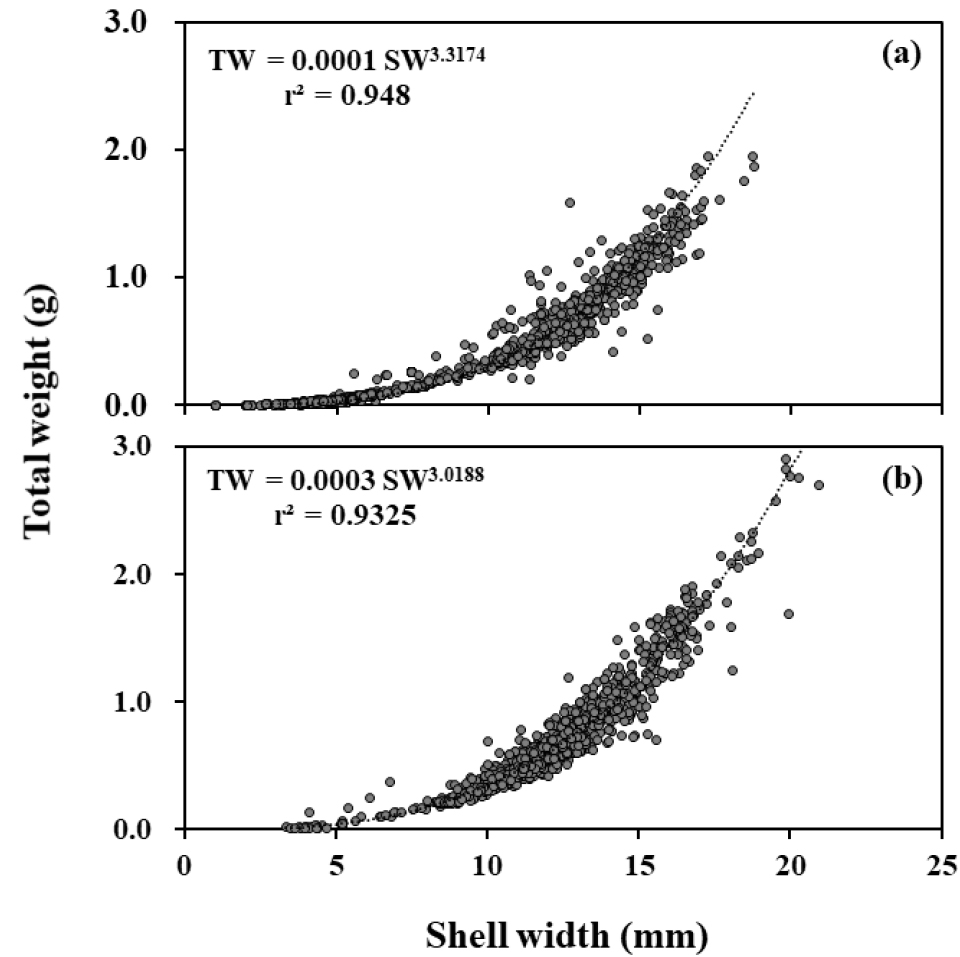
남해안 두 개 하천에서 채집된 기수갈고둥의 각폭과 각고의 상관관계는 봉강천에서 SH = 0.591SW+0.034 (r2 = 0.981), 미룡천에서 SH = 0.637SW-0.359 (r2 = 0.932)로 나타나, 약 10 mm 이하의 각폭일 경우 봉강천의 개체가 각고가 더 높고, 약 10 mm 이상의 각폭에서는 미룡천에 서식하는 개체의 각고가 더 크게 나타났다(Fig. 7). 각폭과 전중량의 상관관계는 봉강천에서 TW = 0.0001SW3.317 (r2 = 0.948), 미룡천에서 TW = 0.0003SW3.019 (r2 = 0.933)로, 미룡천에 서식하고 있는 개체의 무게가 더 크게 나타났다(Fig. 8).
매월 출현 개체를 대상으로 각폭의 크기를 1 mm 간격으로 출현빈도를 분석하였다(Fig. 9). 봉강천은 2018년 3~8월까지 평균 각폭이 증가하였고, 당년에 가입한 각폭 1~2 mm의 개체가 8월에 10.5% 관찰되기 시작하였으며, 9월에는 각폭 2~3 mm의 개체가 23.0%, 10월에는 각폭 3~4 mm의 개체가 57.4%, 11월에는 각폭 4~5 mm의 개체가 50.0%의 높은 빈도를 차지하였다. 미룡천은 봉강천과 달리 각폭의 평균 크기 변화가 크게 나타나지 않았고, 8월에 각폭 1~2 mm의 개체가 확인되지 않았다. 그러나 11월에 출현빈도는 10% 이하로 낮으나 봉강천과 유사한 크기(각폭 3~4 mm)의 개체가 확인되었기 때문에 8월에 어린 개체가 가입되었을 것으로 추정된다.
기수갈고둥의 밀도와 환경요인과의 관계
기수갈고둥 서식 하천에서 정점별 수온 및 염분, 엽록소 a, 부착조류의 총 현존량 및 분류군별 현존량의 환경조건과 기수갈고둥의 서식밀도, 각폭 및 전중량과의 상관관계를 분석하였다. 봉강천에서 수온은 엽록소-a와 출현밀도 및 부착조류 분류군과 뚜렷한 음의 상관관계를 보였고, 각폭 및 전중량과는 높은 양의 상관관계를 보였다. 출현밀도는 엽록소-a와 뚜렷한 양의 상관관계를 보였으며, 부착조류 분류군 중 Melosiraceae, Naviculaceae와 높은 양의 상관관계를 나타냈다(Table 4). 미룡천에서 수온은 엽록소-a와 뚜렷한 음의 상관관계를 보였고, 기수갈고둥의 출현밀도는 염분, 엽록소 a, 부착조류의 현존량, 분류군 중 Bacillariaceae와 뚜렷한 양의 상관관계를 보였다(Table 5).
Table 4.
Correlation coefficients between environmental factors and biotic measurements on Clithon retropictum collected in Bonggang Stream
| Tem. | Sal. | Chl.-a | Abun. | Ba. | En. | Fr. | Me. | Na. | Os. | Density | SW | |
| Sal. | 0.240 | |||||||||||
| Chl.-a | -.629 | -0.188 | ||||||||||
| Abun. | 0.258 | .462 | -0.225 | |||||||||
| Ba. | -.399 | -0.162 | .571 | -0.237 | ||||||||
| En. | -0.398 | -0.285 | 0.177 | -0.293 | .779 | |||||||
| Fr. | -.447 | -0.208 | .656 | -0.150 | .602 | 0.220 | ||||||
| Me. | -.439 | -0.116 | .615 | -0.098 | 0.208 | -0.095 | -0.023 | |||||
| Na. | -.475 | -0.186 | .857 | -0.181 | .687 | 0.302 | .763 | .422 | ||||
| Os. | -.512 | -0.144 | .349 | 0.124 | 0.097 | -0.062 | 0.117 | .437 | 0.179 | |||
| Density | -.417 | -0.188 | .620 | -0.031 | 0.282 | 0.039 | .348 | .739 | .617 | 0.038 | ||
| SW | .693 | .339 | -.559 | .307 | -0.252 | -0.292 | -.448 | -0.374 | -.479 | -0.286 | -.580 | |
| TW | .751 | .324 | -.528 | .415 | -0.299 | -0.331 | -.434 | -.431 | -.464 | -0.267 | -.552 | .946 |
Table 5.
Correlation analysis between environmental factors and biotic measurements on Clithon retropictum collected in Miryong Stream
| Tem. | Sal. | Chl-a | Abun. | Ba. | Fr. | Me. | Na. | Os. | Rh. | Density | SW | |
| Sal | .347 | |||||||||||
| Chl-a | -.463 | -0.084 | ||||||||||
| Abun. | -0.022 | 0.170 | .360 | |||||||||
| Ba. | -.347 | -0.027 | .838 | .405 | ||||||||
| Fr. | -.372 | -0.105 | .562 | 0.280 | .335 | |||||||
| Me. | -.367 | -0.098 | .319 | 0.129 | 0.124 | .735 | ||||||
| Na. | -0.267 | 0.001 | .614 | 0.277 | .858 | 0.148 | 0.054 | |||||
| Os. | -0.118 | -0.085 | 0.108 | -0.081 | 0.093 | 0.015 | 0.075 | -0.009 | ||||
| Rh. | -.388 | -0.098 | .771 | .333 | .540 | .853 | .753 | 0.253 | 0.031 | |||
| Density | 0.112 | .422 | .490 | .517 | .544 | 0.153 | 0.115 | .391 | -0.015 | .383 | ||
| SW | -0.212 | -0.106 | -0.19 | -0.134 | -0.153 | -0.107 | -0.131 | -0.097 | 0.114 | -0.227 | -0.284 | |
| TW | -0.234 | -0.119 | -0.167 | -0.111 | -0.129 | -0.113 | -0.13 | -0.079 | 0.157 | -0.22 | -0.257 | .940 |
*Temp.: water temperature, Sal.: salinity, Chl.-a: Chlorophyll a, Abun.: abundance of epilithic algae, Ba: Bacillariaceae, En: Entomoneidaceae, Fr: Fragilariaceae, Me: Melosiraceae, Na: Naviculaceae, Os: Oscillatoriaceae, Rh: Rhoicospheniaceae, Density: abundance of Clithon retropictum, SW: shell width, TW: total weight
4. 고 찰
기수갈고둥이 서식하는 봉강천과 미룡천은 8월 고고조시 최대 수심이 1.5 m 이내이며, 자갈이나 작은 돌이 풍부한 기수지역으로 수온은 간조시 기온 및 담수의 영향과 만조시 해수 수온에 영향을 받았다. 각 정점에서 수온 변화는 봉강천 6.4~27.0°C, 미룡천 4.7~34.3°C의 범위로, 채집 당일의 측정 시각에 따른 노출 및 기온 변화에 영향을 받았을 것으로 판단된다. 또한 기수갈고둥은 염분변화에 대한 생존능력이 뛰어난데, 5~26 psu의 염분에 서식하고(Kumazawa et al. 1991), 성체는 완전 담수에서 89일간 생존한다(Furujo and Tomiyama 2000). 기수갈고둥이 발견되는 최상부 지점(정점 1)의 연간 염분변화는 봉강천에서 0.05~0.11 psu, 미룡천에서 0.06~0.15 psu의 범위로 나타났고, 서식지의 최하단부에 해당하는 가장 아래쪽 지점(정점 4)의 연간 염분 변화는 봉강천에서 0.08~17.66 psu, 미룡천에서 0.18~33.87 psu로 나타나, 간조시 하천수에 의한 영향과 만조시 해수에 의한 영향을 받았다. 또한 8월 백중사리에 수온, 염분 및 수위 변화에서 확인된 바와 같이, 해당 지역은 조석에 따라 해수의 영향을 크게 받으며 장시간 영향을 미치는 것으로 판단된다. 따라서 기수갈고둥은 완전 담수나 해수에서는 장기간 생존이 불가능하고, 일시적인 상태의 담수 및 해수 조건에서 생존이 가능하며, 기수의 환경조건에서 서식할 수 있는 것으로 판단된다.
고둥류는 치설을 이용해 해조류나 미세조류 등을 섭식하는 초식성과 조개류와 같은 동물의 생체나 사체를 섭식하는 육식성 및 부식성의 특성을 보인다. 기수갈고둥은 초식성으로 기질에 부착한 생물을 섭식하는데, 위 내용물 중에 조류가 있음을 현광현미경을 통해 확인된 바가 있어(Ohara and Tomiyama 2000), 본 연구에서는 부착조류에 대한 분석을 수행하였다. 기수갈고둥의 먹이인 부착조류는 두 하천 모두 유사하게 나타났는데, 겨울철과 봄철에는 규조류가, 여름철과 가을철에는 남조류와 녹조류가 우점함으로써 시기적으로 천이됨을 확인하였다. 하천 및 호소의 식물플랑크톤 군집은 수온이 낮은 시기에는 규조류가, 수온이 증가하는 시기에는 남조류와 녹조류가 우점하는 현상(Reynolds 1984)과 일치하였다. 정점에 따라서도 다소 상이한 분포 특성을 보였는데, 염분이 낮은 지점은 남조류가, 염분이 상대적으로 높은 지점은 규조류가 우점하는 특성을 보였다. 또한 엽록소 a는 수온이 낮은 겨울철에 상대적으로 높은 값을 나타냈는데, 부착조류의 생리적인 특성과 낮은 수온으로 인한 기수갈고둥과 같은 포식 동물의 섭식량 감소로 판단된다.
두 하천 모두에서 기수갈고둥의 출현밀도는 정점 및 시기에 따라 차이가 크게 나타났다. 특히 봉강천에서 가을철에 높은 출현밀도는 산란을 통해 새로 가입한 작은 개체의 출현에 기인한 것으로 평균 각폭 및 중량으로 추정해 볼 수 있고, 겨울철(1~2월)에 두 지역 모두에서 높은 서식밀도는 특정 기질(상대적으로 크기가 큰 돌의 아래)에 고밀도로 서식하였기 때문으로 판단된다. 또한 기수갈고둥은 수온이 적당한 시기에는 먹이나 짝을 찾기 위해 이동하여 넓게 분포하고(Hirata et al. 1992), 수온이 낮은 시기에는 기질 속에 잠입하여 생활하고 있음을 본 연구에서 관찰하였다.
기수갈고둥의 각폭과 각고와의 상관관계는 두 지역이 다소 상이하게 나타났고, 측정된 대부분의 개체에서 각정 부위가 깨져있는 것을 보고하였는데(Noseworthy et al. 2012), 본 연구에서도 이를 확인하였다. 기수갈고둥은 유속이 빠르고 돌맹이가 풍부한 하천에 서식하기 때문에 일시적인 강한 유속으로 부착기질로부터 떨어지거나 움직이는 돌에 의해 패각이 깨졌을 것으로 판단된다. 이는 같은 각폭일 경우, 봉강천에 서식하는 개체가 미룡천에 서식하는 개체보다 각고가 더 낮은 것과 관련이 있으며, 상대적으로 봉강천의 유속이 빠른 것(미발표 자료)과 일치한다. 따라서 각폭과 각고의 상관관계는 사육실험을 통해 기수갈고둥 고유의 성장 패턴을 검증할 필요가 있다.
봉강천에서 기수갈고둥의 산란 시기는 4월부터 7월 하순이었음을 조직학적 방법으로 추정하였다(조와 황 2010). 본 연구에서 봉강천은 3월에, 미룡천은 4월에 처음으로 소수의 난낭(egg capsule)을 관찰하였고, 이후 지속적으로 증가하다가 8월 이후 감소하는 경향을 관찰하였다. 특히 난낭은 기수갈고둥이 서식하는 모든 정점에서 관찰되었다. 기수갈고둥이 법정보호종이기 때문에 해부를 통한 육중량을 측정할 수 없어 각폭에 대한 전중량의 비율로 계산된 비만도(condition factor)가 8월에 소폭 감소하는 것으로 판단할 때 남해안에서 산란기는 3~4월부터 8월로 추정된다. 일본 시즈오카현 이즈반도의 나카강(Naka River, 34°45'N, 138°46'E) 하구에서 기수갈고둥의 난낭은 2월부터 9월까지 관찰되고 2회의 피크를 확인했으며, 부화까지 소요되는 기간은 1개월 이상으로 추정하였다(Nishiwaki et al. 1991b).
기수갈고둥은 피면자 유생으로 부화한 후 부유생활을 하며, 이후 치패로 변태하여 바닥에 착저하게 된다. 일본 오사카현의 칸자키강(Kanzaki River, 34°41'N, 135°25'E) 하구 연구를 통해 어린개체는 조간대 하부 지역에서 주로 8월과 9월 사이에 발생하여 1년 후에 약 9 mm까지 성장한다고 보고하였다(Miyajima and Wada 2014). 또한 후쿠오카현 사이고강(Saigo River, 33°45'N, 130°28'E) 하류에서 8 mm보다 작은 크기의 어린 개체가 기수역 상류 한계점에서 연중 높은 밀도로 출현하며, 담수 지역에서는 발견되지 않았다고 보고하였다(Kobayashi and Iwasaki 2002). 본 연구 지역 중 봉강천에서도 8월 조사에서 각폭 1~2 mm 크기의 개체가 출현하였고, 이듬해 2월에 각폭 약 5 mm까지의 성장을 빈도분포 분석을 통해 확인하였다. 또한 이들 군집은 주로 기수 한계점에 근접한 정점 2에서부터 하천 종단부인 정점 4까지 높은 밀도로 출현하여 어린 개체는 해수의 영향을 크게 받는 지역에 분포하는 것으로 판단된다. 이를 근거로 기수갈고둥의 최초 산란 시기를 3~4월로 가정하고, 1 mm 이하의 어린 개체가 8월에 확인됨에 따라 기수갈고둥의 난발생, 부화, 부유유생, 착저까지의 소요 기간은 수온 및 먹이생물 등 여러 환경요인에 따라 다르지만 최소 60일에서 최대 120일 것으로 판단된다.
돗토리현 하시즈강(Hashizu River 35°29'N, 133°53'E) 하구에서 관찰된 기수갈고둥은 기수역보다 상류의 담수 지역에서 더 큰 개체가 관찰되었고(Kumazawa et al. 1991), 와카야마현 타카세강(Takase River, 33°38'N, 135°24'E) 하구에서는 암컷이 수컷보다 수명이 더 길고, 상류에 서식하는 개체의 연령이 더 높다고 보고하였다(Shigemiya and Kato 2001). 국내 남해안 연초천(이 등 2018)과 본 연구의 봉강천과 미룡천 역시 서식 가능 수역 내에서 상류 지역이 하류 지역보다 크기가 더 큰 개체의 비율이 높았다. 따라서 바다와 연결된 아래쪽 지역에 착저한 어린 개체는 점차 성장하면서 상류 지역으로 이동하는 것으로 판단되나, 향후 국내 하천 기수역에서 기수갈고둥의 난낭 밀도 및 치패 가입량을 포함한 이동 특성에 관한 연구가 필요하다.
기수갈고둥의 분포 특성과 환경요인과의 상관관계 분석에서 서식 밀도와 염분과의 관계를 보면, 봉강천은 상관성이 없었던 반면 미룡천은 양의 상관성을 보임으로써 다른 양상을 나타냈다. 본 연구는 봉강천과 미룡천에서 기수갈고둥이 서식하는 범위 내에 정점을 선정하였고, 채집 및 염분 측정 시각이 조위에 따라 매회 상이하였기 때문에 기수갈고둥의 서식 밀도와 염분과의 상관관계를 특정하기 어려웠던 것으로 판단된다. 향후 기수갈고둥이 서식하지 않는 상류의 담수지역 및 하류의 해수지역을 포함한 넓은 범위에서 간조시 일정 시각이 경과된 후 모니터링하는 연구가 필요하다. 또한 두 지역 모두에서 엽록소 a 농도가 높을수록 양의 상관성(p < 0.01)을 보였다. 어린 개체 분포와 먹이원과 관련하여, 두 지역은 7월부터 11월까지 남조류가 우점하고, 그 외의 시기에는 규조류가 우점하는 특징을 보였다. 따라서, 8월 이후 가입되는 어린 개체는 규조류에 비해 세포벽이 약한 남조류인 Oscillatoriaceae를 초기 먹이원으로 소비하고, 일정 크기까지 성장 후에는 규조류 Bacillariaceae를 주요 먹이원으로 선호할 것으로 판단된다. 본 연구에서 확보된 기수갈고둥 자연개체군 모니터링 자료는 종 복원을 위한 실내증식의 기초 자료, 특히 먹이원 선택을 위해 매우 중요하다고 판단된다.
