

1. 서 론
2. 재료 및 방법
연구해역
원핵생물 개체수 및 생산력
원핵생물 다양성
기타 분석
3. 결 과
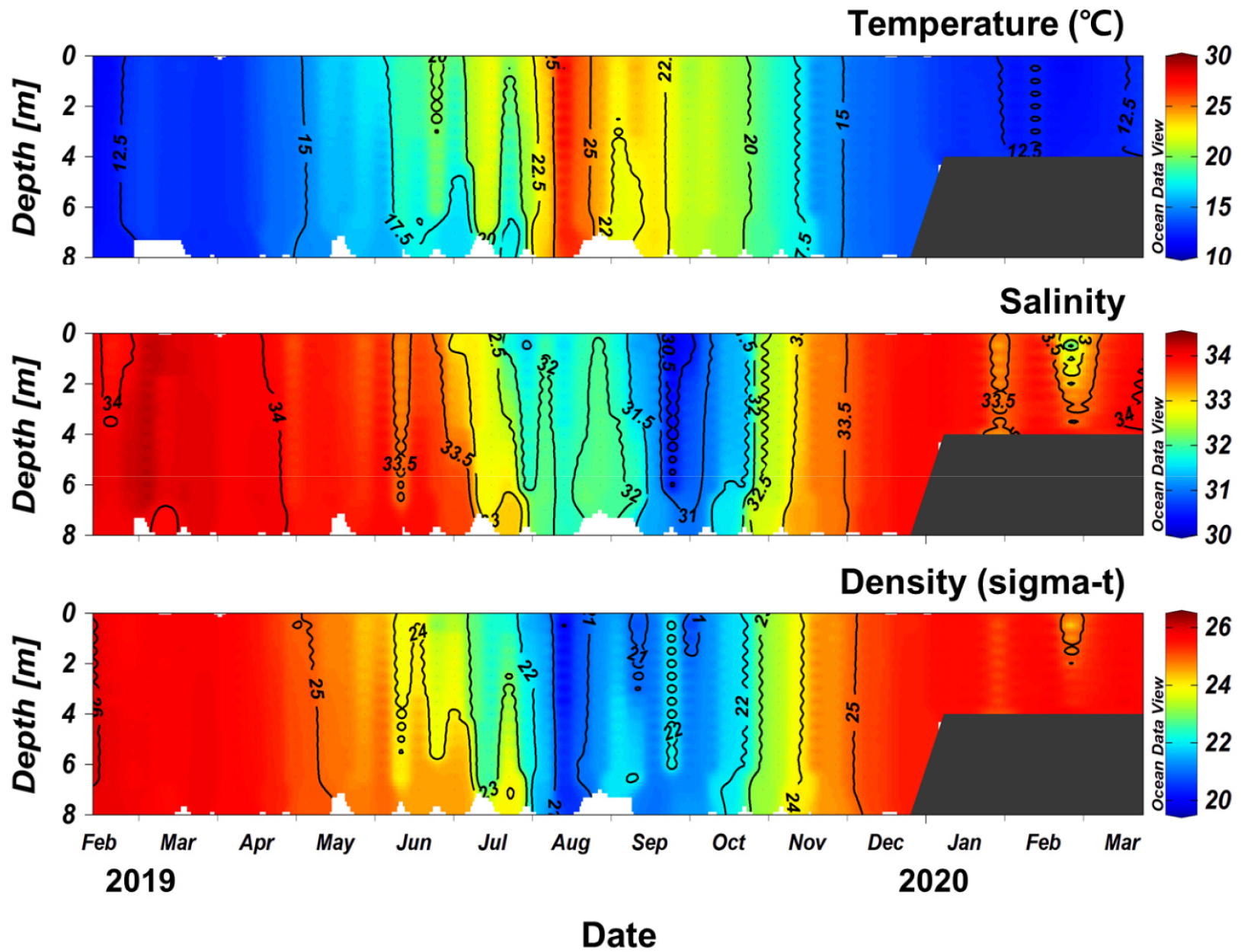
해역의 환경 특성
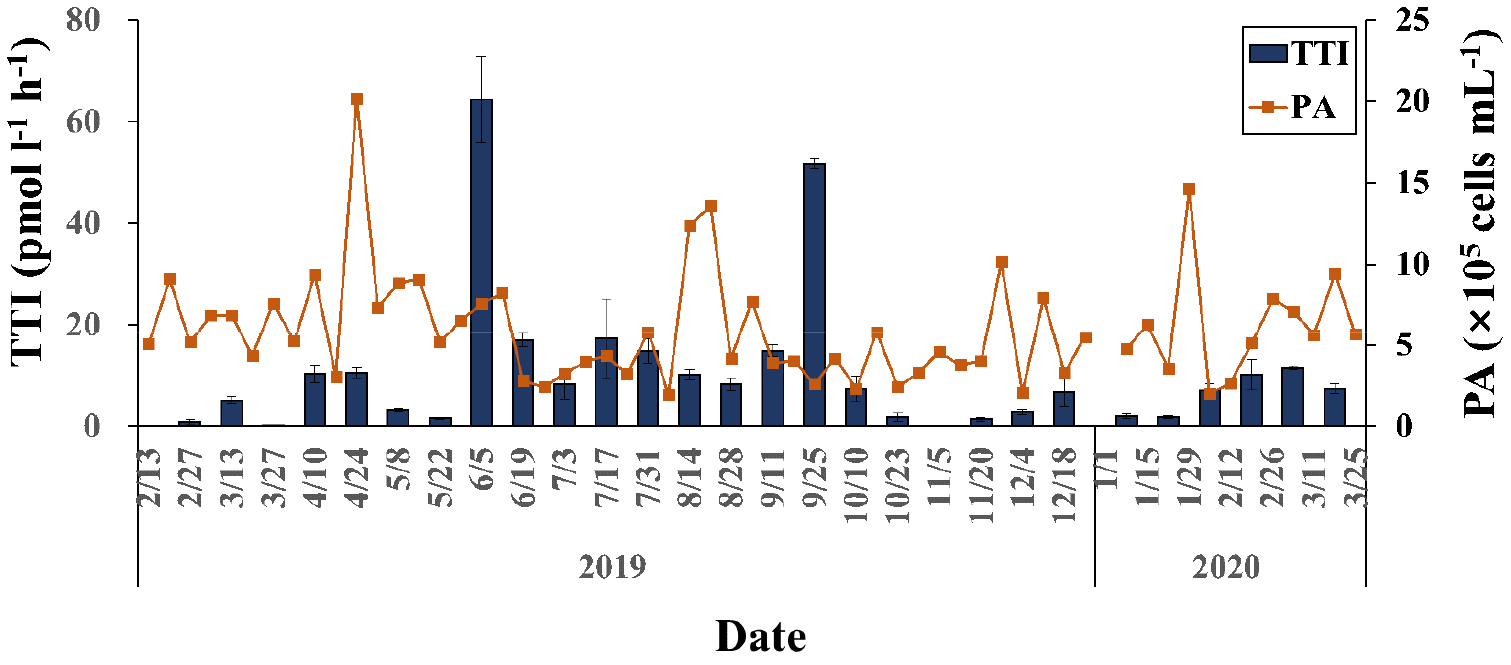
원핵생물 개체수 및 생산력
원핵생물 다양성
4. 고 찰
원핵생물 개체수 및 생산력의 연변동
원핵생물 종조성의 연변동
1. 서 론
지구표면의 70%를 차지하는 해양은 전지구적 일차생산의 50%를 담당하며 막대한 양의 탄소를 저장하는 저장고로서 인류의 식량 공급 및 지구 기후 조절에 매우 중요한 역할을 수행하고 있다(Falkowski et al. 1998). 광합성을 하는 남세균(cyanobacteria)을 제외한 대부분의 종속영양성 해양 원핵생물은 일차생산자가 생산하는 유기물이나 외부로부터 유입되는 유기물을 이용하여 성장하고 미생물 먹이망을 통해 에너지를 전달하는 핵심적 역할을 수행함으로써 탄소 순환에 매우 중요한 역할을 수행한다(Azamet al. 1983; Gasol et al. 2008). 또한 다양한 유기물을 산화시켜 무기영양염으로 재생산함으로써 영양염(질소, 인 등)을 공급하는 등 다양한 원소의 생지화학적 순환에 결정적인 역할을 한다(Arrigo 2005; Monticelli et al. 2014). 해양에서 이들 원핵생물의 성장은 주로 수온 및 식물플랑크톤이 생성하는 유기물 농도에 의존하는 것으로 알려져 있다(Shiah and Ducklow 1994).
20세기 이후 급격한 기후 변화는 해양 온난화와 산성화를 야기하는 등 생태환경을 변화시키고 있다. 북서태평양의 경계에 접한 한반도 주변 해역은 전세계에서 가장 빠르게 수온이 증가하여 기후변화의 영향에 매우 민감한 해역으로 평가된다(Belkin 2009). 이러한 해양 환경의 변화는 궁극적으로 해양 생물의 다양성, 기능 및 생산성의 변동을 야기할 뿐만 아니라 다양한 생물 상호작용의 변화, 즉 먹이망 구조를 변화시킴으로써 머지않아 생태계가 지구환경 및 인류에 미치는 다양한 서비스 기능의 변동이 나타날 것으로 예측된다. 따라서, 이러한 해양환경 및 생태계의 변화를 추적하고 그 변화를 예측하는 것은 지속 가능한 생태계의 이용 및 기후변화에 따른 경제 및 산업적 대응 체계의 구축에 매우 중요하다.
기후 변화에 따른 온난화와 성층 강화로 인한 영양염 감소는 해양 생태계를 지탱하는 식물플랑크톤의 군집 조성에 영향을 미치고 이들의 변화는 연쇄적인 먹이사슬로 연결되어 있는 생물군의 변화를 야기할 것이다. 또한 연안 환경에서는 강 및 하천에서 유기물의 유입, 재부유 등으로 인한 많은 영양염의 유입으로 인하여 해역의 식물플랑크톤 및 연관된 생물군의 군집 조성의 변동이 나타난다(Crossland et al. 2005). 따라서 여러 군집의 일 변동, 계절 변동 및 연 변동 특성과 상호변동성에 대한 이해는 환경변화에 따른 해양생태계의 변동을 세부적으로 이해하는데 중요하다. 그러나 현재까지 대부분의 연구가 계절 변동 또는 단기적 변동에 제한되어 수행되었으며(Alonso-Sáez et al. 2007; Kim et al. 2014; Needham and Fuhrman 2016), 연중 변동 특성을 이해하기 위한 연구는 매우 제한적으로 수행되어졌다(Teeling et al. 2012).
본 연구에서는 남동해 연안인 부산 영도 내만의 고정 관측 지점에서 1년 이상 매주 조사를 통해 원핵생물 군집 특성의 시계열적 분포를 연구함으로써 온대 연안 해역 원핵생물의 군집의 계절 변동성을 이해할 뿐만 아니라 환경 변화에 따른 기초 생태계 변동을 파악하기 위한 기반 자료를 확보하고자 하였다.
2. 재료 및 방법
연구해역
본 연구는 2019년 2월 13일부터 2020년 3월 25일까지 주 1회씩 총 58회 진행하였으며, 2019년 2월부터 2019년 12월까지는 부산 영도구의 한국해양대학교 부근 정점(KMOU, Fig. 1), 그리고 2020년 1월부터는 해양박물관 부근 정점(KNMM, Fig. 1)에서 시료를 채취하였다. 두 정점의 수심은 각각 약 7 m와 3 m로 얕았다.
원핵생물 개체수 및 생산력
원핵생물 개체수는 유세포분석기(flowcytometer)를 이용하여 측정하였다(Marie et al. 1997). 채수한 해수 1 mL을 40 µm 나일론 망을 이용하여 여과한 후 여과 해수에 SYBR Green I (1×, Sigma, MO, US)을 넣어 암실에서 약 10분간 염색하였다. 염색이 끝난 후 샘플을 NovoCyte Flowcytometer System (Agilent, California, US)을 이용하여 계수하였다.
원핵생물의 생산력은 [3H-methyl]thymidine 고정법(Choi et al. 2005)을 이용하여 2주 간격으로 측정하였다. 각각 10 mL의 시료 5개에 3H-thymidine (Specific activity 80 Ci mmol-1)을 최종 농도의 10 nM이 되게 첨가한 후 약 2시간 동안 현장 수온 조건에서 암배양 하였다. 이 때, 2개의 튜브에는 포르말린을 최종 농도 2%가 되도록 넣어 배경값으로 이용하였다. 배양이 끝난 후 3개의 시료 튜브에 포르말린을 최종 농도 2%가 되도록 넣어 반응을 중지시킨 후 약 15분간 냉장 보관하였다. 이후 0.2 µm 셀룰로즈에스터 막여과지(A020A025A; Advantec, Tokyo, Japan)에 시료를 여과하였으며, 얼음물로 냉각시킨 3 mL의 5% Trichloroacetic acid (Sigma, MO, US)로 3회, 얼음물로 냉각시킨 80% ethanol (Merck, Darmstadt, Germany)로 2회 여과지를 헹궈주었다. 여과가 끝난 후 여과지를 20 mL 섬광계수병에 담은 후 Ethyl acetate (Sigma)를 1 mL씩 넣어 녹이고, 10 mL의 섬광계수용 칵테일(Ultima gold; PerkinElmer, Oswestry, UK)을 넣은 후 액체섬광계수기(Tri-Carb 2910 TR; PerkinElmer)로 방사능량을 측정하였다.
원핵생물 다양성
원핵생물의 다양성은 16S rRNA 유전자의 염기서열을 분석하여 조사하였다. 채수한 샘플 0.5 L를 0.2 µm Supor® PES 막 여과지(Pall Corporation, NY, US)에 여과하였다. 여과지를 1.5 mL cryovial에 넣고 1 mL의 STE 완충용액(100 mM NaCl, 10 mM Tris-HCl, 1 mM EDTA, pH 8.0)을 넣어준 후, -80°C에 냉동보관 하였다.
DNA 추출을 위해 샘플을 녹인 후, Somerville et al. (1989)의 방법을 참조하여 lysozyme (5 mg/mL; Sigma, MO, US)을 넣고 37°C에서 30분, sodium dodecyl sulfate (Sigma, MO, US) 및 proteinase K (Qiagen, Hilden, Germany)을 넣고 55°C에서 2시간동안 배양하여 미생물 세포를 용해시켰다.
용해된 샘플에 Phenol-chloroform-isoamyl alcohol (25: 24:1; Sigma, MO, US) 0.5 mL 넣고 약 1분간 부드럽게 뒤집어 충분히 필터지를 녹이고 혼합한 후 5분간 13,000 × g로 원심분리하고 상등액 1 mL을 분리하였다. 분리한 상등액에 1 mL의 Chloroform-isoamyl alcohol (Sigma, MO, US)을 넣고 다시 잘 섞고 5분간 13,000 × g로 원심분리한 후 상등액 0.7 mL을 다시 분리하였다. 분리한 샘플에 0.5 mL의 isopropyl alcohol (Sigma, MO, US)과 sodium acetate (Sigma, MO, US) 70 µL를 넣어준 후 서서히 뒤집어 혼합하였다. 이후 시료를 모두 DNA 정제용 spin column (Biofact, Daejeon, Korea)에 넣고 원심분리하여 column에 걸리게 한 후, 각각 AW1과 AW2 용액(Qiagen, Hilden, Germany)을 넣고 원심분리하여 불순물을 제거하였다. 마지막으로 70°C로 덥힌 AE 용액(Qiagen, Hilden, Germany)을 넣고 DNA를 회수하였다.
원핵생물의 16S rRNA 유전자의 V3-V4 (hypervariable) 구역의 증폭을 위하여 Pro341F (5’-CCTACGGGNBGCASCAG-3’) 와 Pro805R 프라이머(5’-GACTACNVGGGTATCTAATCC-3’)를 사용하였으며(Takahashi et al. 2014), Takara Ex Taq (Takara, Shiga, Japan)을 이용하여 증폭시켰다. 증폭된 PCR 산물은 AMPure XP beads (Beckman Coulter, CA, US)를 이용하여 정제하였으며, 이 산물을 이용하여 MiSeq sequencing을 위한 indexing PCR을 진행하였다(Illumina 2013). 두 번째 PCR이 완료된 후 두 번째 정제를 실시하였으며, 정제된 산물은 Nanodrop 1000 spectrophotometer (Thermo Scientific, MA, US)를 이용하여 정량 하였다. 동량의 PCR 산물을 한 튜브에 모은 후 천랩(Seoul, Korea)의 Illumina MiSeq 2 × 300 PE platform을 이용하여 염기서열을 얻었다.
얻어진 염기서열은 Choi et al. (2018)의 방법을 이용하여 분석하였다. 염기서열 분석은 Mothur 프로그램을 이용하였으며(Schloss et al. 2009), 분석 과정은 표준분석프로토콜(http://www.mothur.org/wiki/MiSeq_SOP)을 참조하였다(Kozich et al. 2013). 먼저 양방향에 얻어진 reads로부터 contig를 만든 후, 400 ~ 525 bp 범위의 길이를 가지며, homopolymers가 8 이하인 contigs를 선택하였다. 염기서열 배열(alignment)은 EzBioCloud 16S database (http:// www.ezbiocloud.net/resources/pipelines)를 참고서열로 사용하였으며, 이후 염기서열 간에 최대 4 base의 차이를 허용한 pre.cluster 명령어를 이용하여 PCR 오류를 최소화하였다. chimera 시퀀스는 chimera.vsearch를 이용하여 제거하였으며, cyanobacteria의 16S rRNA 시퀀스를 제거하여 분석을 진행하였다. 97%의 이상의 시퀀스 유사도를 갖는 시퀀스를 하나의 operational taxonomic units (OTUs)로 정의하였으며, 각 OTU는 80% bootstrap 값 이상의 옵션에서 naïve Baysian classifier를 이용하여 계통분류학적 분석을 수행했다(Wang et al. 2007). 각 시료에서 얻어진 염기서열의 개수는 2,173에서 61,350까지로 변이가 컸으며, 본 연구에서는 염기서열 숫자에 따른 시료간 차이를 최소화하기 위해 최소값인 2,173으로 시퀀스를 재추출하여 시료간 reads 수를 동일하게 한 후 이후의 분석을 진행하였다. 본 연구에서 얻어진 시퀀스 정보는 NCBI의 SRA 데이터베이스에 등록(accession no. SRR13369974 ~ SRR13370031)하였다.
기타 분석
수온 및 염분 자료는 Sea-bird 19Plus CTD (Sea-bird Electronics, WA, US)를 이용하여 수직변화를 측정하였다. 해수는 5 L Niskin 채수기를 이용하여 약 1 m 깊이에서 채수하였다. 채수 후 30분 이내에 실험실로 이동하여 각 연구 항목에 맞게 시료를 처리하였다. 엽록소 a 농도는 해수 시료 0.5 L를 GF/F 여과지로 여과한 후 95% acetone으로 하루 정도 추출한 후 형광분석기(10AU fluorometer; Turner Designs, CA, US)를 이용하여 측정하였다(Parsons et al. 1984). 환경과 원핵생물 군집간의 상관관관계를 확인하기 위하여 Minitab version 12를 이용하여 Pearson 상관관계 분석을 진행하였다.
3. 결 과
해역의 환경 특성
수온은 겨울인 2019년 2월에 11.5°C에서 여름인 8월에 27.6°C까지 변화하여 겨울에 낮고 여름에 높은 전형적인 온대 해역의 계절적 패턴을 보였다. 관측 기간 동안의 평균 수온은 16.7°C였다(Fig. 2). 염분은 2019년 2월 34.18에서 9월 30.26까지 감소한 후 2020년 1월에 33.90까지 증가하였으며, 관측 기간 동안의 평균 염분은 33.10로 나타났다. 6 ~ 11월 사이에 수온은 크게 증가하다 감소하였으며, 염분은 반대로 크게 낮아지다 높아지는 반대의 변동을 보였으며, 그 기간을 제외하면 비교적 좁은 범위의 변동이 나타났다. 한편 2020년 2월 강수로 인하여 염분이 낮아지는 등 일시적 변동도 드물게 나타났다. 본 연구 정점에서 계절에 따른 성층이 뚜렷하게 발달하지 못하였는데, 이는 주로 얕은 수심에 기인한 것으로 여겨진다.
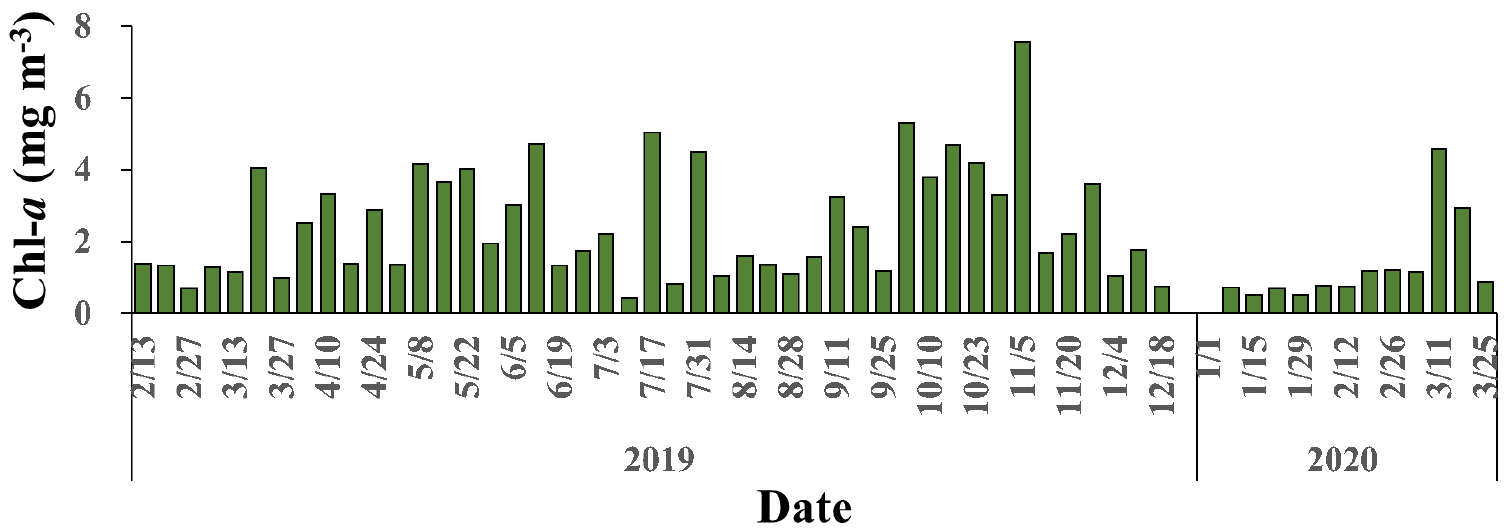
관측 기간 동안 엽록소 a 농도는 0.43 mg m-3에서 7.58 mg m-3까지 나타났으며 시기에 따라 큰 변화를 보였다(Fig. 3). 동계에 상대적으로 낮은 수치를 나타낸 후 봄철 대증식 기간을 거치며 증가하였다. 이후 하계에 접어들며 감소하다 추계에 다시 크게 증가하였으며, 겨울에 다시 낮아졌다. 한편, 하계에는 일시적으로 높은 값이 관측되기도 하였다.
원핵생물 개체수 및 생산력
원핵생물 개체수는 2.0 ~ 20.1 × 105 cells mL-1 범위로 조사기간 동안 약 10 배의 변동을 보였다(Fig. 4). 대체로 봄에 다소 높은 범위의 값을 보였으나 뚜렷한 계절적 양상을 보이지는 않았다. 다만 때때로 10.0 × 105 cells mL-1 이상의 높은 값이 나타나기도 하였다. 원핵생물의 생산력을 나타내는 3H-thymidine 고정율(Thymidine Thymidine Incorporation rates, TTI)은 0.2 ~ 64.3 pmol L-1 h-1 범위로 조사 기간 동안 약 300배의 이상의 큰 범위에서 변동하였다(Fig. 4). 대체로 겨울철인 1 ~ 3월에 0.2 ~ 11.5 pmol L-1 h-1 사이의 낮은 범위의 값을 보였으며, 여름철인 7 ~ 9월 사이에는 8.3 ~ 17.4 pmol L-1 h-1 사이의 비교적 높은 값을 보여 계절적인 변동이 다소 크게 나타났다. 한편, 6월 5일과 9월 25일에는 전후 조사에서 보다 약 6배 높은 TTI가 나타나기도 하였다.
원핵생물 다양성
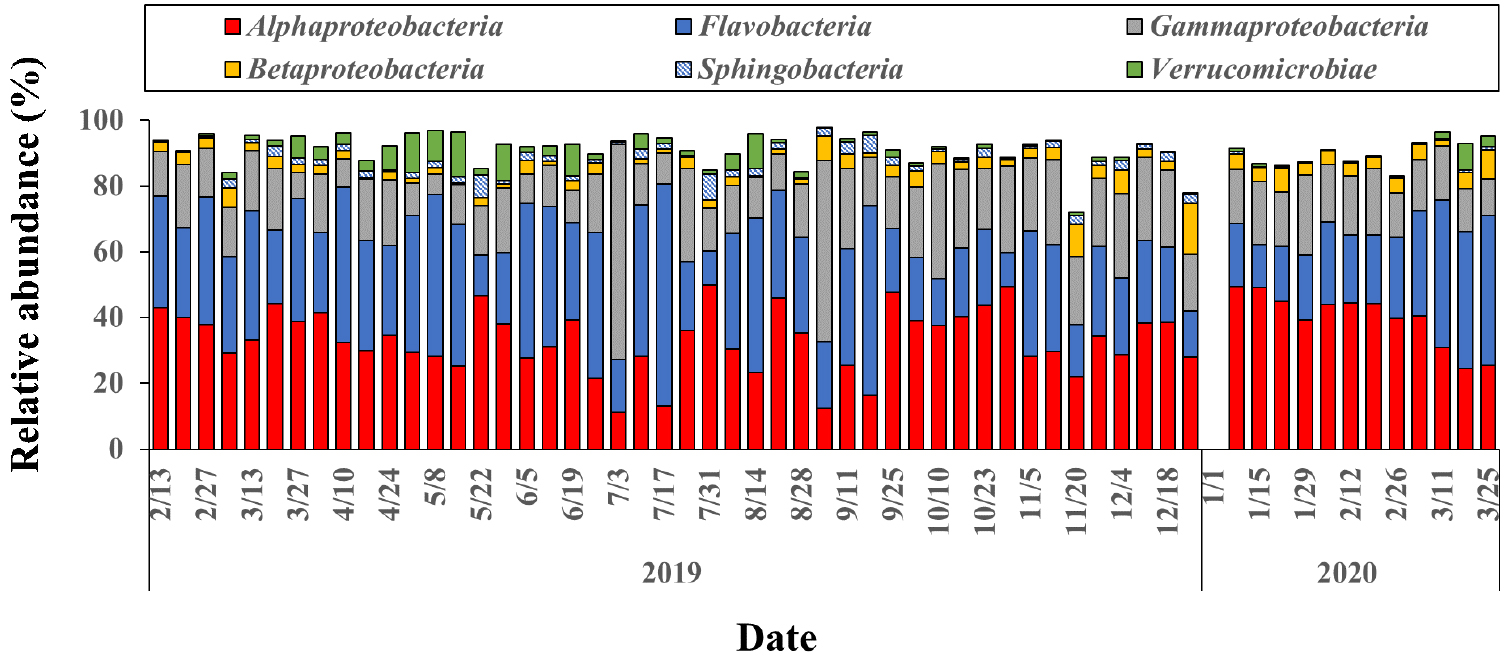
차세대염기서열 분석법을 통해 16S rRNA 유전자의 97% 유사도 기준에서 원핵생물의 OTU는 전 조사 기간에 225 ~ 664 범위의 값을 보여 조사 지점의 원핵생물의 출현종 수는 비교적 작은 변동을 나타내었다. 최소한 한 시료에서 5% 이상 출현하여 우점한 원핵생물의 OTU는 44개로 나타났다. 강 수준에서 볼 때 Alphaproteobacteria가 연평균 34.5±9.7%로 가장 우점하였으며, 뒤를 이어 Flavobacteria가 30.0±12.6%, Gammaproteobacteria가 18.7± 9.8%, Betaproteobacteria가 3.5±2.5%, Verrucomicrobiae가 2.6±3.4%, 그리고 Sphingobacteria가 1.7±1.5%로 나타나 강 수준에서 점유율은 큰 차이를 보였다(Fig. 4).
Alphaproteobacteria는 뚜렷한 계절적인 변동 특성을 나타내지 않고 꾸준히 조사해역에서 대부분의 시기에 가장 우점하는 군집으로 나타났다(Fig 5). 반면 Flavobacteria는 겨울에 낮았다가 봄이 되면서 점차로 증가하는 양상을 보였으며, 여름과 가을에는 크게 증감하는 양상을 나타냈다. 그 밖에 봄에는 Verrucomicrobiae, 봄부터 가을 사이에는 Sphingobacteria, 가을과 겨울에는 Betaproteobacteria가 유의하게 증가하는 계절적 변화 양상을 나타냈다. 7월과 9월 초에 Gammaproteobacteria가 일시적으로 55% 이상 크게 증가하였으며, 이때 Alphaproteobacteria와 Flavobacteria가 20% 이하로 모두 크게 감소하였다.
강 수준에서 큰 변이는 나타나지 않은 반면, 종 수준에서의 연간 변화는 매우 다양하고 큰 변동 양상을 보였다. 특히 가장 우점하였던 Alphaproteobacteria는 종 수준으로 보았을 때 연중 우점종의 변화가 가장 뚜렷하였다(Fig. 6). Pelagibacter가 연중 나타났으나 일정한 시기적 변동 경향 없이 0 ~ 29.6% 범위로 매우 큰 폭의 변이를 나타냈다. Planktomarina는 2월부터 7월초까지 0.02 ~ 18.3% 범위로 이 기간 Pelagibacter와 최우점하는 종으로 나타났지만, 그 이후부터 12월까지 최대 2.8%로 거의 나타나지 않는 뚜렷한 계절성을 나타내었다. 반면, Rhodobacteraceae과에 속하는 AVDB_s OTU는 7월부터 12월에만 나타났으며, 이 기간에 Alphaproteobacteria 중 최우점하였다. 그 밖에 Lentibacter algarum도 대부분의 시기에 출현하였으며 동계에 대체로 10.1%까지 높은 출현율을 나타냈다.
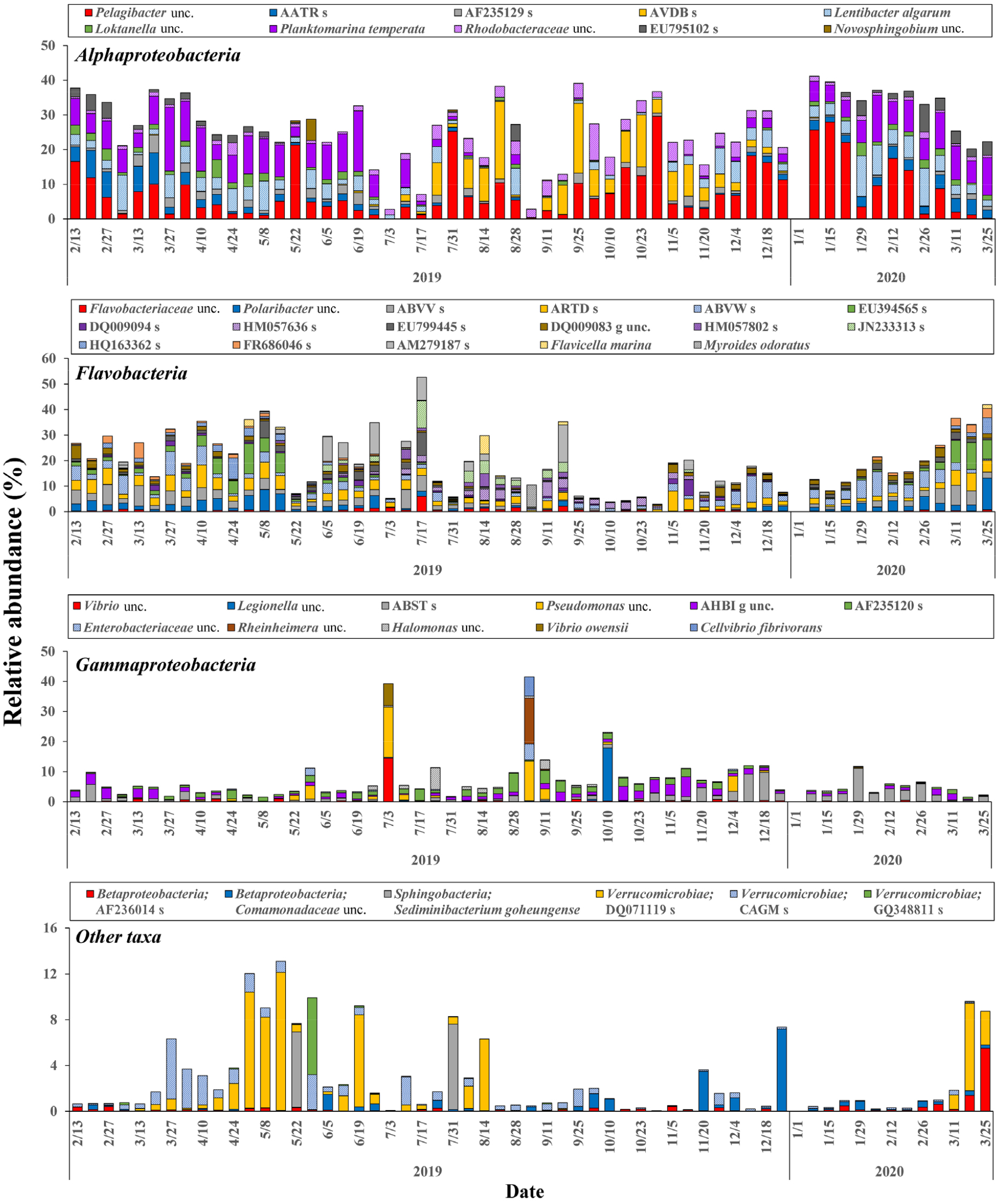
Flavobacteria는 대부분의 시기에 20% 이상 높은 분포 특성을 보였으며, 춘계에 증가하여 최대값에 도달한 후 겨울까지 감소하는 계절 변동 특성을 나타냈다(Fig. 6). Flavobacteria는 적어도 한 조사에서 5% 이상의 출현한 OTU가 17개로 가장 많았는데, 많은 수의 OTU가 큰 점유율의 변동폭을 보이며 연중 나타나는 특성을 보였다. 하지만 Ulvibacter에 속하는 OTU (EU394565)는 봄에만 출현한 반면 두 개의 OTU (JN233313, AM279187)는 6월부터 9월 사이에만 출현하였으며, Polaribacter는 하계에는 나타나지 않아 일부의 OTU는 뚜렷한 계절성을 나타내었다.
Gammaproteobacteria는 대부분의 시기에 15% 이하의 출현율을 보였으며, Flavobacteria가 감소하는 가을에서 초겨울 사이에 다소 높은 점유율을 나타내었다(Fig. 6). 연중 3개의 OTU (ABST, AF235120 및 AHBI)가 비교적 높게 나타났는데, 각각 겨울과 봄 사이 그리고 여름과 가을에 상대적으로 높은 점유율로 천이하는 경향을 보였다. 한편, 세 번의 조사에서는 Gammaproteobacteria가 최우점하였는데, 이때는 Vibrio, Pseudomonas, Enterobacteria 및 Rheinheimera 속의 원핵생물이 기회적으로 크게 우점하는 특성을 나타내었다.
그 외 다른 강에 속하는 주요 박테리아는 연중 낮은 수치를 나타내었으나, 대부분 뚜렷한 계절 변동 특성을 나타냈다(Fig. 6). Verrucomicrobiae에 속하는 CAGM_s, DQ071119_s와 GQ348811_s는 모두 봄철에 높은 비율로 나타났는데, CAGM_s가 초봄에 상대적으로 높은 비율을 보이다 감소할 때 DQ071119_s가 약 10% 이상으로 우점하여 천이하는 양상을 보였다. Betaproteobacteria에 속하는 OTU인 AF236014_s는 3월에 약 5%로 높았으나 대부분의 시기에 1% 미만의 낮은 점유율을 보였으며, Comamondaceae OTU는 초겨울에 약 8%까지 높은 점유율로 나타났다. 5월 및 7월에 기회적으로 한번씩 Sphingobacteria에 속하는 Sediminibacterium goheungense가 일시적으로 약 7%까지 증가하였다. 한편, 본 연구에서 고세균(Archaea)도 나타났으나, 연평균 0.7%로 매우 낮게 출현하였다.
4. 고 찰
일 년여에 걸친 기간 동안 수행된 주간 조사를 통해 원핵생물 군집의 연변동 특성을 면밀히 이해할 수 있었다. 원핵생물 개체수는 특이적으로 높은 수치를 기록한 날이 있었으나, 연중 변화는 크지 않았다. 원핵생물 생산력은 겨울에 낮고 여름에 높은 경향을 보였으나, 수온 및 염분만이 유의하나 낮은 상관관계를 보여 다양한 환경요인들이 원핵생물의 생물량과 성장 조절에 영향을 미칠 가능성을 시사하였다. 한편 원핵생물은 강 수준에서 4개의 그룹만 모든 시료에서 출현하여 우점하였으며, 최소한 한 시기에 5% 이상 출현한 그룹도 6개에 그쳐, 연안 환경에서 소수의 원핵생물 강(class)의 생태적 역할에 중요하게 기능함을 시사하였다. 그러나, 종 수준에서는 40여개의 다른 원핵생물들이 최소 5% 이상의 점유율을 보이며 한 시기 이상 출현하였으며, 많은 원핵생물 종들이 유사한 시기에도 큰 폭의 점유율 변동을 보여 계절 규모의 환경 변화보다 더 짧은 규모의 환경 변화에 따라 다양한 원핵생물 종들이 매우 민감하게 반응하여 상호 경쟁하고 있음을 시사하였다.
원핵생물 개체수 및 생산력의 연변동
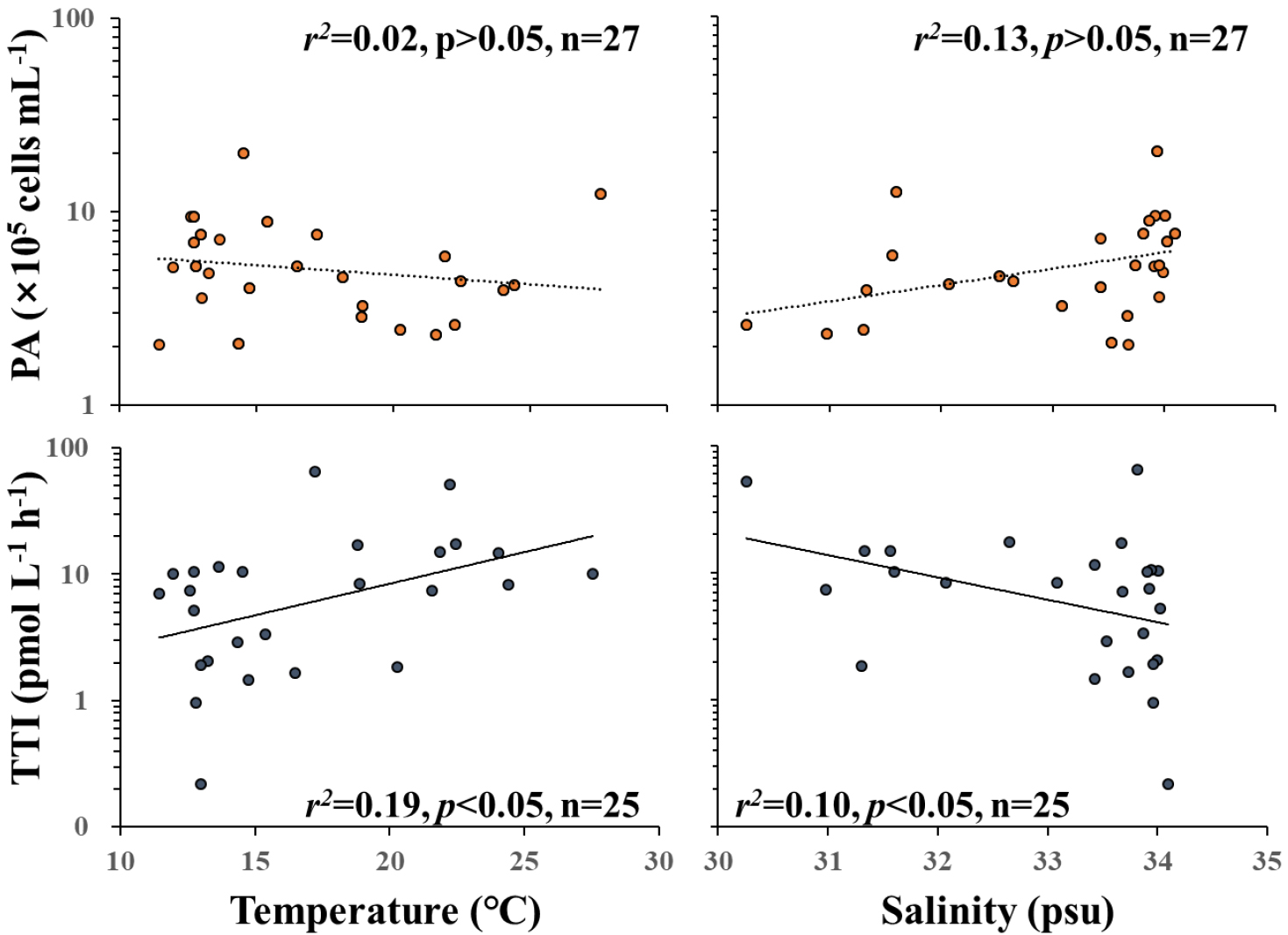
연평균 원핵생물 개체수는 5.7 × 105 cells mL-1로 나타났으며, 연중 계절변화에 따른 개체수의 변화가 크게 나타나지는 않았으나, 상대적으로 봄철 개체수가 다른 계절에 비하면 1.6배 높았다. 이는 과거 봄철 식물플랑크톤 대번성기에 개체수가 증가한다는 연구들과 유사하였다(Hyun and Kim 2003; Sturluson et al. 2008). 원핵생물 개체수는 일반적으로 수온 변화에 따라 양의 변동 특성을 나타내며, 엽록소 a 농도와도 양의 상관관계를 갖는 것으로 알려져있다(White et al. 1991). 그러나 본 연구해역에서는 원핵생물 개체수가 수온 및 엽록소 a의 농도와 유의한 양의 상관관계를 보이지 않아(p > 0.05), 원생생물의 섭식 등 하향식(top-down) 조절 기작에 의해 조절되거나 식물플랑크톤 이외의 유기물 공급원이 있을 가능성을 시사하였으며 이에 대한 연구 필요성을 제시하였다.
원핵생물 생산력도 수온, 엽록소 a 및 원핵생물 개체수와 양의 상관관계가 있음이 알려져 있으며(White et al. 1991), 염분과는 음의 상관관계를 나타낸다고 연구된 바 있다(Ducklow and Kirchman 1983). 또한 과거 Cheaspeake Bay에서 나타난 생산력의 계절별 변화에서 수온이 상승함에 따라 생산력 또한 상승하는 경향이 나타났다(Shiah and Ducklow 1994). 본 연구해역에서도 유사하게 수온과 양의 상관관계, 염분과는 음의 상관관계를 보였으나, 0.19 이하로 낮은 결정계수를 보였는데(Fig. 7), 이러한 낮은 상관성은 본 연구해역에서 원핵생물의 성장이 수온 및 염분 등의 환경 요인에 일부 의존하고는 있으나, 그 영향력이 크지 않음을 시사한다. 뿐만 아니라 용존 유기 탄소(DOC) 농도는 0.74 ~ 3.43 mg C L-1로 나타났으며(미발표 자료), 이는 연구해역에서의 원핵생물 생산력으로 추정된 탄소 요구량의 1,000 ~ 5,000배 이상, 일차생산을 통해 추정된 공급되는 탄소량에 비해서도 5배 이상 높았다(미발표 자료). 따라서 연구해역은 식물플랑크톤 이외의 다양한 유입원으로부터 많은 유기물이 유입되고 있는 것으로 보이며 인접한 소형 하천으로부터의 타지성 유기물 유입 및 인접 저서성 서식환경에 서식하는 미세조류 및 해조류로의 유기물 생산 등이 다른 유기물 공급원들로 생각된다(Kim and Kim 2017; Oh et al. 2017).
원핵생물 종조성의 연변동
연안 해역에서 나타나는 원핵생물 군집 구조는 여러 가지 환경 요인에 의해 변동된다고 알려져 있다(Fuller et al. 2006; Gilbert et al. 2009; Campbell and Kirchman 2013). 본 연구 해역에서 엽록소 a의 영향은 크지 않으나, 수온 및 염분과 같은 환경 요인이 군집 구조 변동에 다소 영향을 주는 것으로 보인다(Table 1). 특히 Alphaproteobacteria 및 Flavobacteria에서 계절 변동이 뚜렷하게 나타났으며, Gammaproteobacteria에서도 작게나마에서도 계절적 변동 양상이 나타났으나, Verrucomicrobiae에서는 계절적 변동 양상이 나타나지 않았다(Fig. 5, Table 1).
Table 1.
Pearson Correlation Coefficient between environmental factors (temperature, salinity, and chl-a) and prokaryotic communities
| Class | Species | Temperature | Salinity | TChl-a |
| Alphaproteobacteria | Pelagibacter unclassified | -0.148 | 0.043 | -0.125 |
| AATR s | -0.613*** | 0.538*** | -0.179 | |
| AF235129 s | -0.088 | 0.134 | 0.07 | |
| AVDB s | 0.599*** | -0.694*** | 0.145 | |
| Lentibacter algarum | -0.501*** | 0.36** | -0.285* | |
| Loktanella unclassified | -0.471*** | 0.414** | -0.214 | |
| Planktomarina temperata | -0.549*** | 0.641*** | -0.191 | |
| Rhodobacteraceae unclassified | 0.474*** | -0.678*** | 0.321* | |
| EU795102 s | -0.453*** | 0.211 | -0.238 | |
| Novosphingobium unclassified | 0.03 | 0.039 | -0.015 | |
| Flavobacteria | DQ009094 s | 0.042 | 0.029 | 0.097 |
| DQ009083 g unclassified | -0.537*** | 0.452*** | -0.115 | |
| EU799445 s | 0.004 | 0.211 | 0.16 | |
| ABVV s | -0.332* | 0.423** | -0.086 | |
| FR686046 s | -0.424** | 0.41** | -0.075 | |
| ABVW s | -0.566*** | 0.386** | -0.249 | |
| ARTD s | -0.315* | 0.466*** | 0.193 | |
| HM057636 s | 0.762*** | -0.713*** | 0.309* | |
| Flavicella marina | 0.316* | -0.157 | -0.074 | |
| Flavobacteriaceae unclassified | 0.393** | -0.175 | 0.143 | |
| HM057802 s | 0.593*** | -0.303* | -0.071 | |
| AM279187 s | 0.376* | -0.123 | 0.136 | |
| JN233313 s | 0.57*** | -0.292* | 0.181 | |
| HQ163362 s | -0.264* | 0.336* | -0.004 | |
| Myroides odoratus | 0.163 | -0.132 | -0.062 | |
| Polaribacter unclassified | -0.403** | 0.425** | -0.121 | |
| EU394565 s | -0.376** | 0.407** | 0.101 | |
| Gammaproteobacteria | AHBI g unclassified | 0.052 | -0.148 | 0.262* |
| Rheinheimera unclassified | 0.184 | -0.16 | -0.052 | |
| ABST s | -0.403** | 0.149 | -0.294* | |
| Cellvibrio fibrivorans | 0.182 | -0.156 | -0.054 | |
| Enterobacteriaceae unclassified | 0.171 | -0.112 | -0.083 | |
| AF235120 s | 0.587*** | -0.456*** | 0.275* | |
| Halomonas unclassified | 0.358** | -0.339* | -0.082 | |
| Legionella unclassified | 0.167 | -0.281** | 0.14 | |
| Pseudomonas unclassified | 0.194 | -0.105 | -0.079 | |
| Vibrio unclassified | 0.061 | -0.008 | -0.014 | |
| Vibrio owensii | 0.083 | -0.021 | -0.015 | |
| Other taxa | Comamonadaceae unclassified | -0.108 | 0.042 | 0.001 |
| AF236014 s | -0.186 | 0.153 | -0.088 | |
| Sediminibacterium goheungense | 0.11 | -0.088 | 0.072 | |
| CAGM s | -0.063 | 0.153 | -0.053 | |
| DQ071119 s | 0.014 | 0.182 | 0.093 | |
| GQ348811 s | 0.015 | 0.066 | -0.024 |
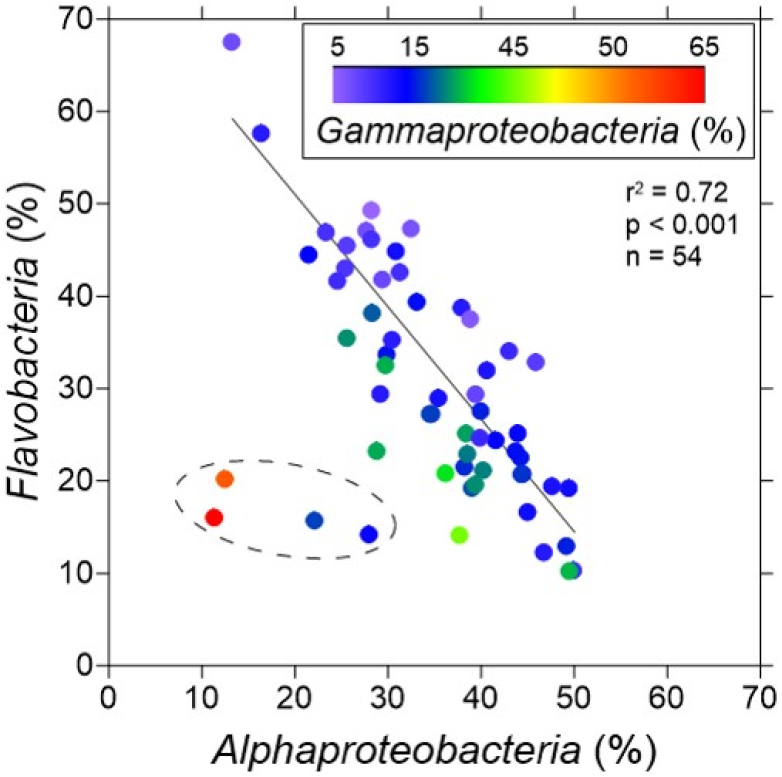
일반적으로 해양에서 우점하는 Proteobacteria와 Bacteriodetes가 본 연구해역에서도 우점하였으나, 강 수준 및 종 수준에서 뚜렷한 차이를 나타냈다. 서영국해협에서의 연구에 의하면, Alphaproteobacteria와 Flavobacteria가 증감하는 양상이 같게 나타나기도 했으나(Gilbert et al. 2009), 본 연구해역에서는 두개의 우점하는 그룹인 Alphaproteobacteria와 Flavobacteria 사이에 반비례하는 분포 특성이 뚜렷하게 나타났다(Fig. 8). 본 연구에서 Alphaproteobacteria와 Flavobacteria는 서로 간에 증감이 반대되는 성향을 보이며, 대부분 약 65%의 우점도를 나눠 가졌다(Fig. 8). 매우 드물게 이 둘의 우점도가 20% 이하로 떨어진 경우가 있었는데, 이때는 예외적으로 Gammaproteobacteria가 50% 이상의 높은 우점도를 보였다.
Alphaproteobacteria중 Pelagibacter는 대부분의 해양 환경에서 높은 비율로 나타나며, 상대적으로 작은 크기를 가져 빈영양 해역에서 특히 우점하는 것으로 알려져 있으다(Straza and Kirchman 2011; Giovannoni 2017). 이들은 본 연구해역에서 약 10% 이하로 낮은 우점도를 보였는데 이는 연구해역이 육상에 인접하여 강물의 유입과 해조류 등 다른 유기물 유입원의 영향을 받아 나타난 결과로 사료된다. 반면 Rhodobacteraceae에 속하는 원핵생물이 우점하였는데, 일반적으로 Rhodobacteraceae가 많이 나타나는 환경은 영양염이 풍부한 부영양 환경으로 알려져(González et al. 2000; Suzuki et al. 2001; Alonso and Pernthaler 2006), 연구 해역이 외부 유기물 등의 유입으로 유기물 농도가 높은 것과 연관된 것으로 보인다. 또한 Rhodobacteraceae는 수온 변화에 따라 군집이 변동하는 특성을 잘 보여주는 그룹 중 하나이다(Selje et al. 2004). 대표적으로 Planktomarina temperata는 15°C의 낮은 수온에 적응하여 살아가는 종으로 알려져 있으며(Giebel et al. 2019), 본 연구해역의 수온이 약 20°C이상 상승하는 7월부터 출현 비율이 급격하게 감소하였다가 다시 수온이 낮아지는 11월경부터 증가하였다(Fig. 6). 이러한 결과는 수온 및 유기물 등 환경 조건에 따라 Rhodobacteraceae내에서 종간의 천이 및 계절적인 군집 조성의 변동 특성이 나타났음을 시사한다.
Flavobacteria는 식물플랑크톤이 대번성하는 봄이나 강 등으로부터 유기물이 유입될 때 증가하는 것으로 알려져 있다(Pernthaler et al. 1998; Mary et al. 2006; Choi et al. 2018). 본 연구해역에서는 연중 Flavobacteria가 꾸준히 나타났으며, 특히 춘계 대번성기가 나타난 봄철에 Alphaproteobacteria가 감소함과 동시에 증가하는 양상을 보였다. 그러나 춘계 대번성기에 나타난 종(Polaribacter, ABVV _s, ABVW_s, EU799445, EU394565 및 FR686046)과 이후 감소하는 시기에 나타난 종(Flavobacteriaceae OTU, HM057636, JN233313 및 AM279187)은 서로 대부분 달랐다. Polaribacter는 낮은 수온에 잘 적응하는 종으로 알려져 있어(Brinkmeyer et al. 2003), 수온의 변화가 Flavobacteria의 계절 변동에 영향을 미쳤을 것으로 여겨지나, 하계에 식물플랑크톤 종조성의 뚜렷한 변이(미발표 자료, 양 등)를 고려하면, 유기물 조건(조성 및 양)의 변화에 따른 Flavobacteria 종조성의 변동 가능성도 배제할 수 없을 것으로 보인다.
Gammaproteobacteria는 연중 낮은 우점도를 보였으나, 세 개의 OTU (ABST, AHBI 및 AF235120)가 연중 우점하면서 계절에 따른 종간 우점도의 천이가 발생하였다. 겨울에 우점하였던 AHBI에서 봄이 되며 AF235120으로 우점군집의 천이가 일어났으며 추계에 ABST로, 겨울에 다시 AHBI가 우점하는 계절적 변동 특성을 보였다. 또한 특이적으로 세번의 조사(7/3, 9/4 및 10/10)에서 Vibrio, Pseudomonas, Enterobacteria 및 Rheinheimera 속이 원핵생물 중 가장 우점하였는데, 본 연구 자료에서는 파악하기 어려운 급격한 환경 변화에 기회적으로 성장하였거나, 어류의 배설물 등 군집 조성의 비 정상적 교란을 야기할 수 있는 유기물 덩어리 등이 시료에 유입된 결과가 아닌가 여겨진다.
그 밖의 그룹은 연간 꾸준히 우점하는 종들이 아닌 특정한 시기에 한시적으로 등장하는 종들이 대부분이었다. 특히 Verrucomicrobiae는 대부분의 경우 낮은 비율로 출현한 원핵생물 그룹 중 하나이나, 식물플랑크톤 생체량과 큰 연관이 있으며, 봄철 대번성기가 진행되며 많이 나타나는 군집으로 알려져 있다(Lindh et al. 2015; Bunse et al. 2016). 본 연구에서도유사하게 Verrucomicrobiae는 춘계에 대번성기가 진행되며 급격하게 증가한 후 대번성기가 끝나며 감소하는 경향을 보였다(Fig. 5).
한편 2019년의 4 ~ 5월경에서 나타난 봄철의 군집 구조의 변화가 2020년에는 3월경부터 발생하여 약 한달 정도 앞서 나타났다. 이는 2020년 2월의 평균 수온이 2019년 2월의 평균 수온보다 약 0.3°C 높게 나타났으며, 질산염, 암모늄, 인산염 및 규산염이 각각 1.6배, 1.7배, 1.2배 및 1.4배로 높았는데(미발표 자료), 높은 영양염 농도 및 다소 증가된 수온 등 환경 조건의 변화가 이러한 원핵생물 군집의 출현 시기에 영향을 미쳤을 가능성이 있을 것으로 보이나, 이러한 변화 요인을 이해하기 위해서는 더 긴 장기 조사가 필요할 것으로 여겨진다.
요약하면, 매주 조사를 통해 원핵생물 군집의 생물량, 생산성 및 군집 조성의 연변동 특성을 이해할 수 있었다. 생물량은 비교적 좁은 범위에서 연간 변동하였으나, 생산력은 약한 계절성을 나타내며 큰 폭의 변동을 보였다. 반면, OTU 수준에서의 군집 조성은 종류에 따라 다양한 시기적 변동 양상을 나타내어 원핵생물의 생태적 기능 또한 크게 변동될 가능성을 시사하였다. 장기적 연구를 통해 이들 군집 조성의 변동을 조절하는 요인에 대한 이해 및 지구온난화와 같은 환경 변화에 따른 연안생태계의 역할과 기능의 변동에 대한 이해가 가능할 것으로 기대된다.
