

1. 서 론
2. 재료 및 방법
현장조사
Chlorophyll-a 농도와 영양염 측정
수온과 염분도 연직 분포 및 성층화 지수(Stratification Index)
식물플랑크톤 정량 및 정성 분석
T-S 다이어그램 및 통계분석
3. 결 과
표층 환경요인
수온과 염분도 연직 분포 특성
식물플랑크톤 현존량과 우점 및 아우점 속
T-S 다이어그램 및 통계분석
4. 토 의
2022 춘계에서 하계 제주 연안 표층수의 물리화학적 변동
여름철 양자강 희석수 유입과 제주 연안 성층화에 미치는 영향
식물플랑크톤의 현존량 및 종조성 특징
5. 요약 및 결론
1. 서 론
식물플랑크톤은 해양 생태계의 주요 일차생산자로서, 먹이 사슬의 핵심 역할을 수행한다. 식물플랑크톤의 군집 구조는 수온, 염분, 영양염의 농도 등 다양한 해양 환경 요인의 변화에 따라 변동할 수 있으며, 먹이 사슬 상위 영양 단계 생물의 군집 구조 및 생산력에 영향을 미친다(Joo et al. 2016; 박과 이 2017). 이러한 해양 환경요인은 주로 계절 또는 공간적 특성에 따라 변동하게 되는데, 춘계 온대해역에서는 일반적으로 겨울철 수괴의 혼합으로 인한 풍부한 저층 영양염이 표층으로 공급되며 일조량이 증가함에 따라 식물플랑크톤의 대발생이 나타나지만(Carstensen and Conley 2004), 하계에는 표층 수온 상승으로 인한 성층화 현상으로 인해 저층의 영양염이 표층으로 공급되지 않아 식물플랑크톤 성장이 저해될 수 있다(Riley 1942; Cushing 1989; 이 등 2016). 한편, 공간적 관점에서 연안 해역은 용승현상 및 육상 기원의 영양염류 유입으로 인한 해양 물리화학적 환경변화가 식물플랑크톤의 높은 생산성을 유도한다(손과 백 2015). 그러므로 온대 연안생태계의 식물플랑크톤 군집 변동은 시간적/공간적 해양환경의 복합적 변화에 따라 나타난다. 기후 변화의 최전선에 위치한 제주 연안은 지리적 특성으로 인해 계절에 따라 대마 난류(Tsushima Warm Current), 제주 난류(Jeju Warm Current), 양자강 희석수(Changjiang River Diluted Water) 등 다양한 해류 및 수괴에 의해 물리화학적 변화가 크게 나타난다. 양자강 희석수의 경우, 강우량이 많은 여름철 양자강 댐의 방류로 인해 대규모의 저염분 수괴가 제주도 남서쪽 해역으로 유입되는 것으로 알려져 있다(Park 1986; 김 등 2019). 양자강 희석수의 높은 수온과 낮은 염분 및 높은 영양염 함량은 제주 연안 생태계에 다양한 영향을 미칠 수 있는 것으로 알려져 있다(김 등 1998; Kim et al. 2013; Affan et al. 2014). 최근 기후변화로 야기된 여름철 빈번한 폭우 및 홍수 등으로 인한 양자강 희석수 수괴의 범위 확장 및 표층 수온의 증가는 제주 연안의 식물플랑크톤 군집 등 전반적인 제주 연안 생태계에 직접적인 영향을 줄 수 있을 것으로 판단되지만(Oh et al. 2023), 이와 관련된 체계적이고 장기적인 조사는 현재 이루어지고 있지 않다. 이전 제주 연안의 식물플랑크톤 군집 구조에 관한 연구는 주로 바다 목장이나 항만 부근 등 특수 지역을 대상으로 이루어져 왔다(이 1989; 이 등 1998; 신과 이 2002; 윤 2015, 2016). 2015년 제주 연안 전역을 대상으로 식물플랑크톤 군집 구조를 모니터링한 이전 연구의 경우, 제주 연안의 환경요인에 따른 식물플랑크톤 주요종의 출현에 대한 분석이 이루어졌으나, 영양염을 비롯한 화학적 환경요인을 고려하지 않았으며, 표층 및 수심 30 m 의 수온과 염분, 용존산소만 모니터링하였기 때문에 온대 해역 식물플랑크톤 군집에 중요한 영향을 미치는 성층화 현상 및 수괴 특성 등의 물리적 환경요인 역시 제대로 고려되지 못했다(김 등 2019). 또한 해당 조사는 9년 전 진행된 것이며, 최근 제주 해역에서 평균 수온 상승으로 열대 기원 산호의 서식지 범위가 점차 확대되는 등 기후변화로 인한 해양환경 변화를 고려하였을 때 지속적인 조사가 필요하다(Kim and Kang 2022; Yang et al. 2023). 따라서, 본 연구는 2022년 제주 연안의 영양염, 성층 구조 및 수괴 특성 등 해양 물리화학적 환경 요인 변화에 따른 식물플랑크톤 종들의 출현 특성을 확인하며, 장기적으로는 제주 연안 생태계의 기초적 자료를 수집하고 기후변화 등 해양 환경 변화에 대응하기 위한 장기 모니터링 기반을 확립하고자 하였다.
2. 재료 및 방법
현장조사
제주 연안의 물리화학적 특성 및 식물플랑크톤의 출현 변동을 파악하기 위해 2022년 식물플랑크톤의 현존량이 높을 것으로 기대되는 봄철부터 여름철인 3월부터 8월까지 제주 인근 해역 동, 서, 남, 북 총 12정점(J1–J12)을 대상으로 제주대학교 조사선(R/V)인 ‘제라호’를 활용하여 조사를 수행하였다(Fig. 1, Supplementary Table 1). 각 정점의 선상에서 다항목수질측정기(Professional Plus, YSI incorporated, USA)를 사용하여 표층의 해수 수온과 염분도를 측정하였고, 제주 연안의 성층화 현상 추적을 위해 CTD (SBE 911 plus, Sea-Bird, USA)를 사용하여 연직 수온과 염분도 분포 데이터를 기록하였다. 식물플랑크톤의 정량, 정성 분석, 클로로필 농도(Chlorophyll-a, Chl.-a), 영양염(질산염+아질산염; NO3+NO2, 인산염; PO4, 규산염; SiO2) 농도의 측정을 위해 각 정점에서 채수기(10L PVC Niskin, General Oceanics, USA)를 이용하여 수심 1 m에서 해수를 채수하였다.

Chlorophyll-a 농도와 영양염 측정
클로로필(Chl.-a) 및 영양염 농도 측정을 위해 해수 1 L를 직경 47 mm의 GF/F 여과지(Glass Microfiber Filters, Whatman, Ireland)로 여과하였다. 여과지는 Chl.-a 농도 측정을 위해 물기를 제거한 후 빛을 차단하여 영하 80°C에서 냉동 보관하였고, 여과된 해수 역시 영양염 농도 측정을 위해 10 mL 튜브(conical tube, SPL, Korea)에 넣어 분석 전까지 냉동 보관하였다. 실험실에서 Chl.-a 농도 측정을 위해 냉동 보관한 여과지를 실온에서 해동한 후 90% acetone 10 mL를 첨가한 뒤 냉암소에서 24시간 동안 클로로필을 추출하였다. 추출된 Chl.-a 의 농도는 분광 광도계(Epoch2, Agilent, USA)에서 파장 750, 665, 645 및 630 nm의 흡광도를 측정하여 계산 및 보정하였다(Parsons et al. 1984). 영양염 농도 측정을 위해 냉동 보관한 시료를 실온에서 해동한 후 규산염, 인산염, 질산염+아질산염의 농도를 Rho et al. (2015)의 분석방법에 따라 영양염 자동분석기(QuAAtro, SEAL Analytical, Germany)를 이용하여 측정 및 계산하였다. 각각의 영양염 농도는 분석방법의 정확도와 정밀도를 확인하기 위해 표준물질(RMNS, KANSO Technos Co., Ltd., Japan Chemical Industries, Japan)을 함께 사용하여 보정하였다(노 등 2015).
수온과 염분도 연직 분포 및 성층화 지수(Stratification Index)
선상에서 획득한 제주 연안 12개 정점의 수직 수온과 염분도 CTD 기록은 후보정하여 수심 1 m 간격으로 측정값을 산출하였다. 산출된 결과는 Surfer software ver. 21 (Golden Software, USA)을 사용하여 시각화한 후 조사 기간 동안의 성층화 정도를 파악하였으며, 조사 시기 및 정점 별 성층화 정도를 다음 식을 활용하여 성층화 지수(Stratification Index, SI)로 산출하였다(Simpson et al. 1978).
위 식에서, h는 수심(m)을 나타내며, 𝜌는 밀도(kg/m3), 는 밀도의 연직 평균, g는 중력가속도(m/sec2), z는 표층부터 저층까지 연직방향으로 측정된 데이터 간격을 의미한다. SI = 0일 때 수괴는 수직적으로 완벽히 혼합된 상태이며, 성층의 안정도가 증가할수록 SI 값은 증가한다.
식물플랑크톤 정량 및 정성 분석
식물플랑크톤의 정량, 정성분석을 위해 해수 중 1 L를 불투명한 광구병에 담고 루골용액(Lugol’s solution, Sournia 1978)을 넣어 최종 농도 1%가 되도록 현장에서 즉시 고정하였다. 실험실로 옮겨진 고정된 시료는 24시간 이상 정체시킨 후, 상층액을 제거하여 100 mL로 농축하였고 Sedgewick-Rafter Chamber에 분주하여 광학현미경(BX50, Olympus, Japan)을 사용하여 200배 및 400배 배율에서 동정 및 계수하였다. 20 µm 이하의 광학현미경 상에서 동정이 어려운 편모류는 기타 편모조류(unidentified flagellates)로, 편모가 관찰되지 않은 종들은 미동정 나노플랑크톤(unidentified nanoplankton) 수준으로 분류하였다. 계수된 자료는 L당 세포수로 환산하였으며, 총 현존량에서 높은 비율을 차지한 속을 순차적으로 우점속과 아우점속으로 기록하였다.
T-S 다이어그램 및 통계분석
조사 기간 동안 제주 연안에서 관측된 식물플랑크톤의 우점 및 아우점속의 출현 특성을 파악하기 위해 각 조사 시기 및 조사 정점에서 10% 이상의 점유율을 차지한 우점속, 아우점속을 T-S 다이어그램(T-S diagram) 상에 표기하였다. 수온 12°C 이상, 염분도 33.5–34 psu 범위를 가지는 제주난류 수괴와 수온 14°C 이상, 염분도 34.1 psu 이상의 범위를 가지는 대마난류 수괴, 수온 23°C 이상 염분도 31 psu 이하의 양자강 희석수 수괴의 정보를 사전 연구를 참조하여 T-S 다이어그램에 표기하였다(Gong et al. 1996; Lie et al. 2001; KOSEF 1990). 조사 기간 동안 클로로필 농도와 식물플랑크톤의 현존량과의 상관관계 확인을 위해 Pearson 상관분석을 SPSS Statistics ver. 24(IBM, USA)를 활용하여 수행하였다. 또한, 측정된 제주 연안의 물리화학적 환경요인 특성과 식물플랑크톤 출현과의 상관성을 이해하기 위해 RDA (redundancy analysis)를 R software ver. 4.3.1을 활용하여 수행하였다.
3. 결 과
표층 환경요인
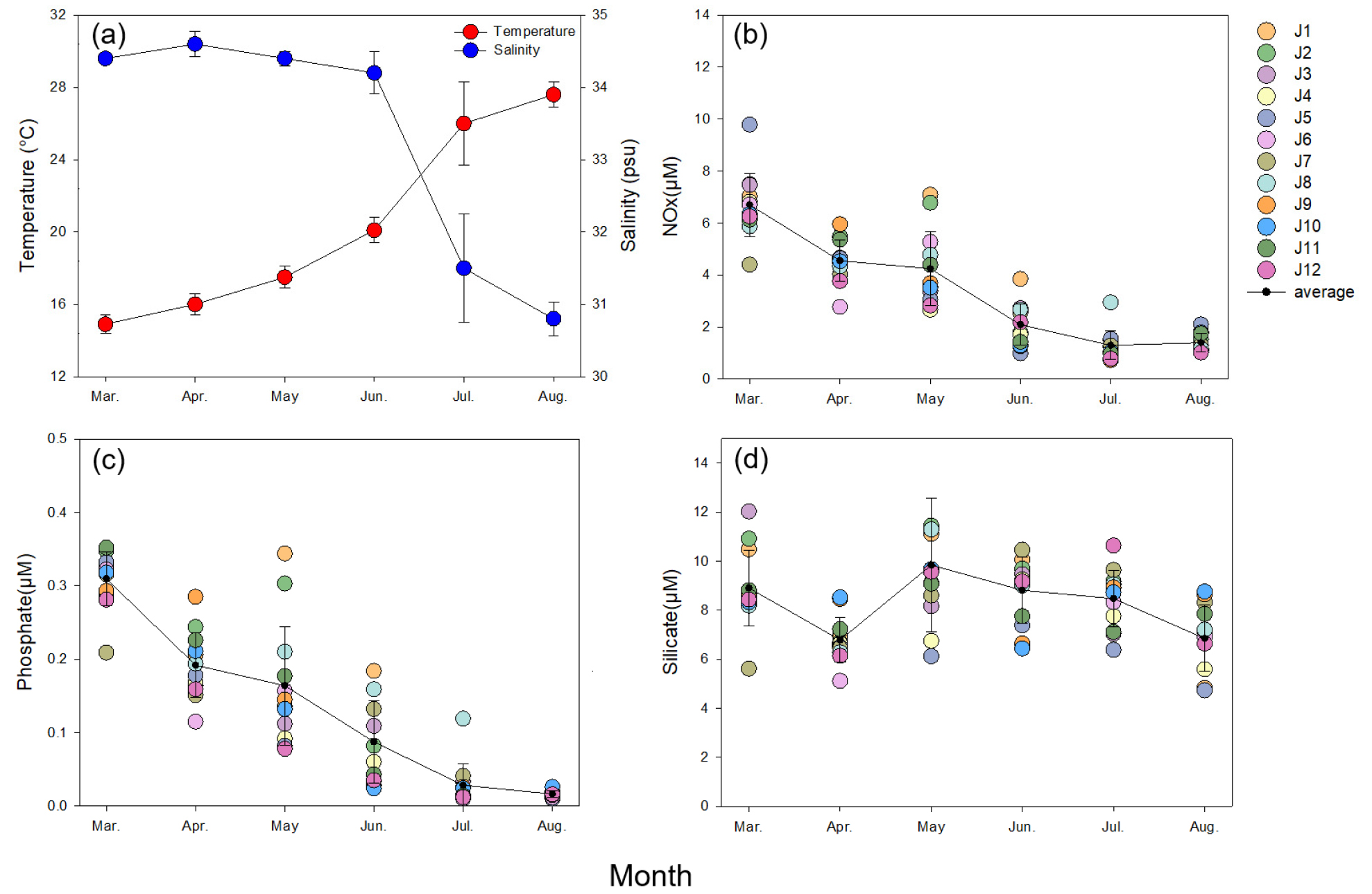
조사기간 동안 제주 연안 12개 정점의 평균 표층 수온은 월별 14.9–27.6°C의 범위로 관측되었으며, 가장 낮은 수온 값을 보인 3월부터 증가하기 시작하여 8월에 수온이 가장 높은 전형적인 온대 해역의 특징을 보였다(Fig. 2a). 염분도는 30.8–34.6 psu의 범위로 관측되었는데, 4월에 가장 높고, 이후 감소하기 시작하여 8월에 가장 낮은 값을 나타내어 수온과 반대 경향을 보였다(Fig. 2a). 영양염의 경우, 질산염+아질산염의 농도는 월별 평균 1.3–6.7 µM의 범위로 측정되었으며, 3월에 가장 높은 농도를 보였다가 하계로 갈수록 점차 감소하는 경향을 보였고 7월에 가장 낮은 농도를 나타냈다(Fig. 2b). 인산염은 월별 평균 0.02–0.31 µM의 범위로 측정되었고, 질산염+아질산염 농도와 동일하게 3월에 가장 높은 농도를 보였다가 하계로 갈수록 점차 감소하여, 8월에 가장 낮은 농도를 나타냈다(Fig. 2c). 규산염은 월별 평균 6.8–9.8 µM의 범위로 측정되었으며, 5월에 가장 높았고 4월에 가장 낮게 측정되었다(Fig. 2d). 질산염+아질산염과 인산염의 농도는 조사 기간 내내 지속적으로 감소하는 경향을 보인 반면, 상대적으로 규산염의 농도는 조사 기간 동안 일정 범위(6.8–9.8 µM) 내에서 변동하였다. 각 영양염 항목을 정점 별로 비교하였을 때, 조사 기간 동안 영양염의 농도가 다른 정점에 비하여 유의미하게 높거나 낮은 정점은 구분되지 않았다(Fig. 2b–d).
수온과 염분도 연직 분포 특성
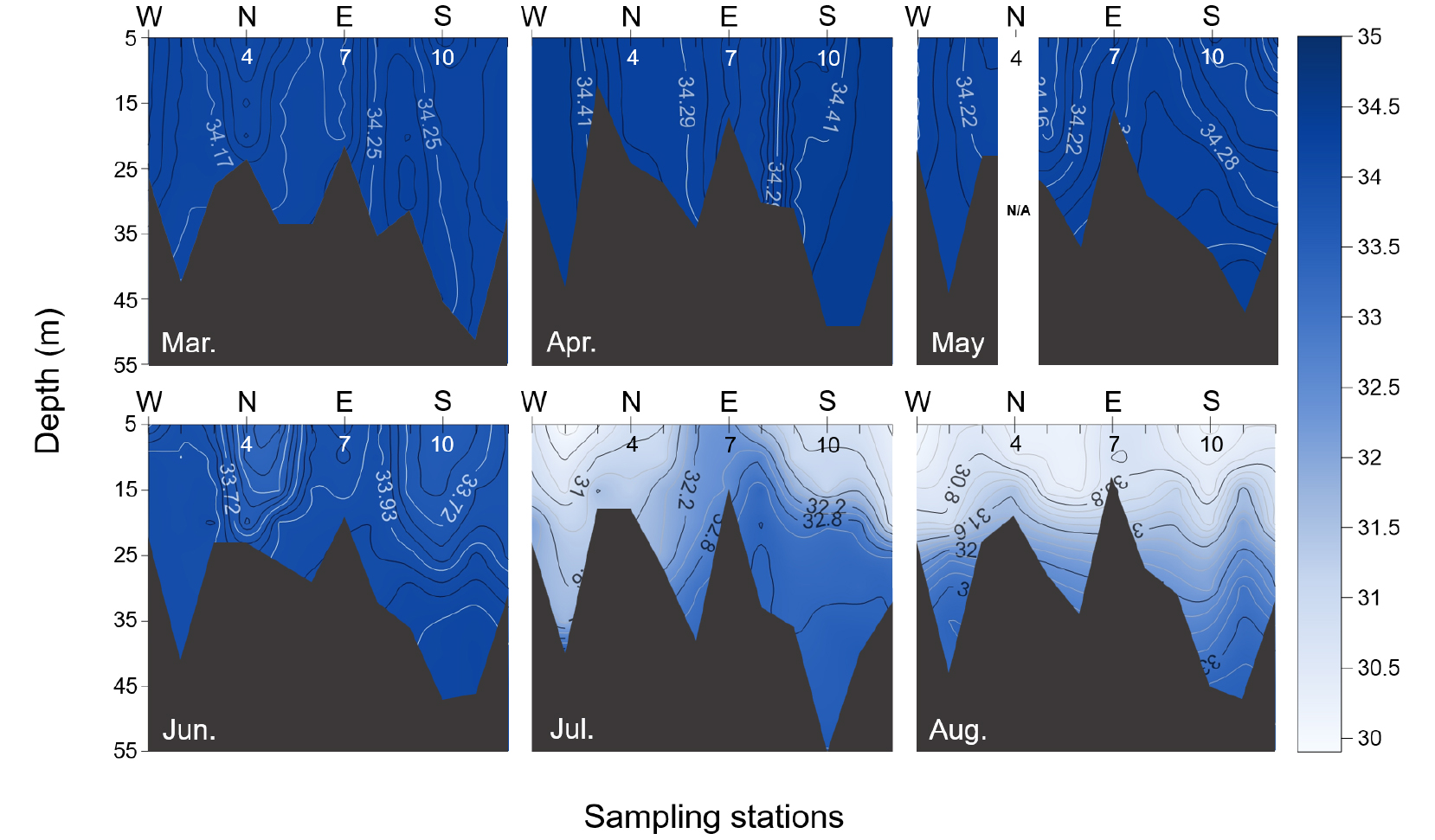
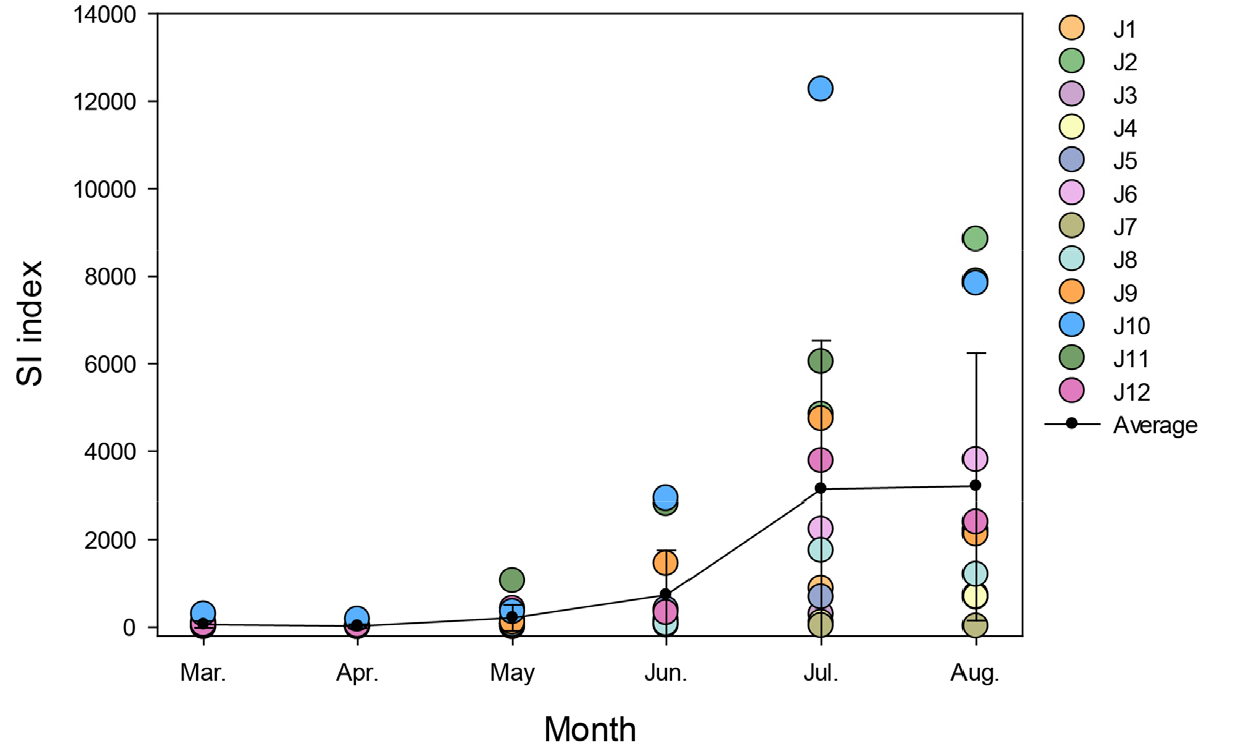
조사 기간 동안 제주 연안의 성층화 현상을 관찰하기 위해 수온과 염분도의 수직 분포도를 작성하였다(Figs. 3 and 4). 3–5월은 성층이 형성되지 않고 수괴가 잘 혼합되어 있었으며, 하계가 시작되는 6월부터 표층 수온의 증가로 약한 성층이 형성되기 시작하였다. 7월에는 고온・저염의 표층수가 북동쪽을 제외한 대부분의 정점에 분포하였으며, 하계가 시작되는 6월부터 표층 수온의 증가로 약한 성층이 형성되기 시작하였다. 해당 시기에 수온이 가장 높고 염분도가 가장 낮았던 정점은 제주 서쪽 지역의 J2 정점(27.9°C, 29.9 psu)으로서, 수온이 가장 낮았던 J7 정점과의 표층 수온의 차이는 6.6°C, 염분도 차이는 2.4 psu로 나타났다. 8월에는 7월보다 고온·저염의 표층수가 제주 연안 전역에 고르게 분포되었으며, 가장 고온 및 저염분을 기록한 지역은 서쪽의 J1 정점으로 각각 29.3°C, 30.1 psu으로 측정되었다. 7월 대비 최고 수온은 1.4°C 상승하였으나, 최저 염분도는 비교적 일정하게 유지되었다. 또한, 모든 조사 정점의 수심 15–25 m 부근에서 수온 및 염분약층이 강하게 형성되어 8월의 제주 연안은 7월보다 성층화 된 것을 확인할 수 있었다. 성층화 지수(SI) 계산 결과, SI 값은 월별 평균 22–3,196 J/m3 범위로 나타났으며, 수층 혼합이 잘 이루어진 3–5월에는 비교적 낮은 평균 SI 값을 나타내었으나, 6월부터 상승하기 시작한 후 7월부터 급격히 상승하여 8월까지 높은 값을 유지하였다(Fig. 5). 다만, 13–18 m의 얕은 수심을 가진 정점들(J1, J3, J4, J7)은 7, 8월 임에도 불구하고 표층과 저층의 수온 및 염분도 차이가 크지 않아 하계임에도 불구하고 비교적 낮은 SI값을 나타내었다(Figs 3 and 4).
식물플랑크톤 현존량과 우점 및 아우점 속
조사기간 동안 관찰된 식물플랑크톤의 현존량은 14,323–2,065,882 cells L-1의 범위로 나타났다(Fig. 6). 12개 정점의 평균 식물플랑크톤의 현존량은 4월과 5월 각각 406,053, 409,139 cells L-1로 가장 높게 나타났으며, 최고 현존량은 4월 J6 정점에서 2,065,882 cells L-1으로 기록되었다. 해당 기간 동안 정점 간의 현존량의 차이는 비교적 컸으며, 조사 시기에 따라 차이가 있지만 주로 제주 북쪽 및 동쪽에 위치한 정점들(J3–J9)에서 높은 경향을 보였다(Fig. 6a). 성층의 발달이 뚜렷하게 관찰되었던 6월부터 8월에는 월별 평균 45,546–118,214 cells L-1 범위로 춘계에 비해 비교적 낮은 식물플랑크톤 현존량을 보였다. 식물플랑크톤의 현존량을 유추할 수 있는 Chl.-a 의 농도는 0.3–1.8 µg L-1의 범위로 나타났는데 7월을 제외한 월 평균값은 대체적으로 식물플랑크톤의 현존량 값과 유의미하게 통일된 변동패턴을 보여주었다(r = 0.949, p < 0.05)(Fig. 6b). 7월의 경우 식물플랑크톤의 현존량이 낮았으나, Chl.-a 의 농도는 평균 1.8 µg L-1로 조사 기간 중 가장 높은 Chl.-a 농도값을 기록하였다.
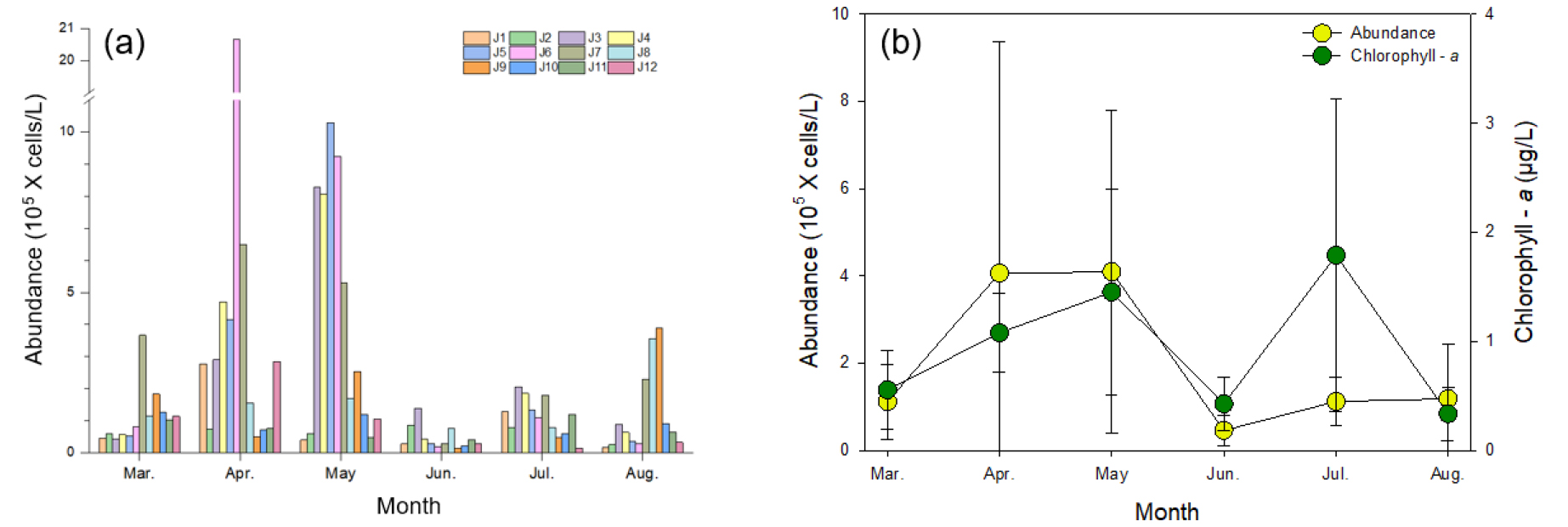
조사 기간 동안 관찰된 식물플랑크톤은 총 47속 84종이 동정되었으며, 분류군 수준으로 살펴보면, 규조류(Bacillariophyta)가 전체 평균 70.1%로 가장 많이 출현하였고, 기타 편모조류(unidentified flagellates) 11.2%, 와편모조류(Dinophyta) 11.0%의 순으로 규조류의 출현 비율이 높았다(Fig. 7). 조사 시기별로 살펴보면, 춘계에 해당하는 3월, 4월, 5월 동안 규조류는 각각 81.7%, 79.0%, 68.9%, 미동정 나노플랑크톤(unidentified nanoplankton) 9.0%, 9.8%, 6.7%, 기타 편모조류가 0.1%, 7.2%, 14.3%의 비율로 출현하였으며, 성층의 발달이 관찰된 6월, 7월, 8월에는 규조류 61.7%, 72.6%, 56.7%, 와편모조류 10.9%, 10.2%, 23.7%, 기타편모조류 17.3%, 12.1%, 16.1% 순으로 출현하여 하계로 갈수록 편모조류의 출현이 증가하는 것을 확인할 수 있었다. 각 조사 시기 및 정점 마다 가장 높은 점유율을 차지한 상위 두 개의 속을 우점속 및 아우점속으로 기록하였다(Table 1). 조사 기간 동안 규조류 4속(Chaetoceros, Skeletonema, Pseudo-nitzschia, Cylindrotheca), 와편모조류 1속(Heterocapsa)으로 총 5속이 우점속으로 출현하였고, 그 외 규조류 9속, 와편모조류 2속 그리고 유글레나류 1속이 아우점속으로 출현하였다. 특히, 조사기간 동안 와편모조류인 Heterocapsa 속은 본격적으로 성층이 발달하는 6월부터 10% 이상 우점도를 보이는 등 눈에 띄게 출현 비율이 증가하는 경향을 보였다.
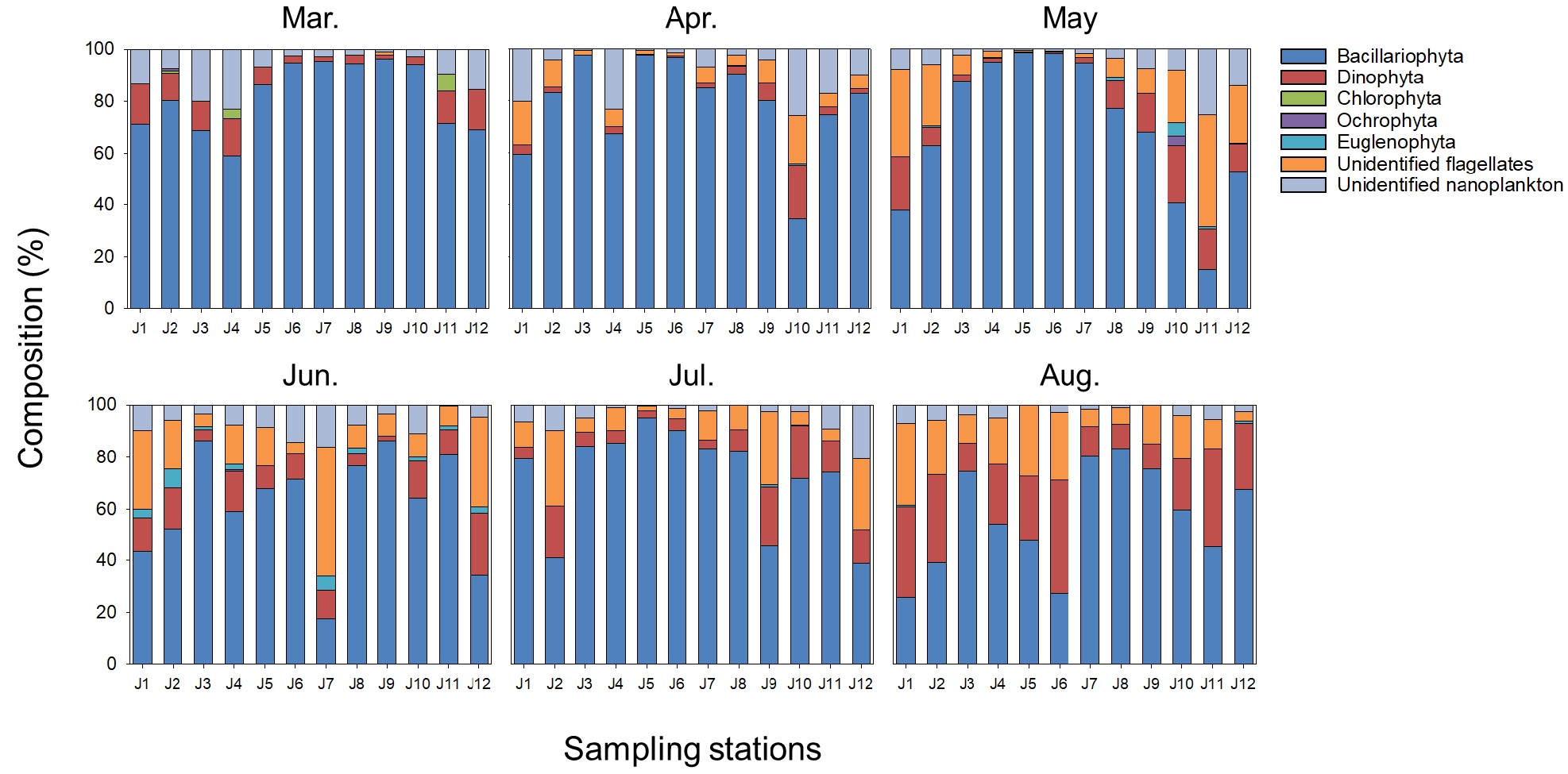
Table 1.
Dominant and subdominant genus at each station during study period. Each color of background means; Yellow: < 10%, orange: 10% ≤ and < 40%, red: 40% ≤
T-S 다이어그램 및 통계분석
T-S 다이어그램을 이용한 분석결과, 제주 연안은 3–5월에는 대마난류 수괴, 6월에는 북쪽의 일부 정점을 제외하고 제주난류 수괴의 물리적 특성에 해당하였다. 해당 기간 동안 물리적 특성이 유사한 두 수괴의 영향을 받은 제주 연안은, 일부 정점에서 와편모조류인 Heterocapsa가 우점 및 아우점 속으로 출현하였으나 대부분의 정점에서 Chaetoceros와 Skeletonema 등의 규조류가 우점하는 분포 특성을 보였다(Fig. 8, Table 1). 그리고 7월 8월 하계 제주 연안은 7월 동쪽의 일부 정점들을 제외하고는 염분도 31 psu 이하, 수온 23°C 이상의 양자강 희석수 수괴의 물리적 특성을 나타내었고, 해당 기간 동안 규조류인 Chaetoceros의 출현이 상대적으로 줄어들었으며 Bacteriastrum과 Cylindrotheca,Pseudo-nitzschia 등 다양한 규조류가 아우점속으로 분포하였고, 와편모조류인 Heterocapsa와 Gyrodinium이 빈번하게 10% 이상의 점유율을 차지하는 아우점속으로 출현하였다.
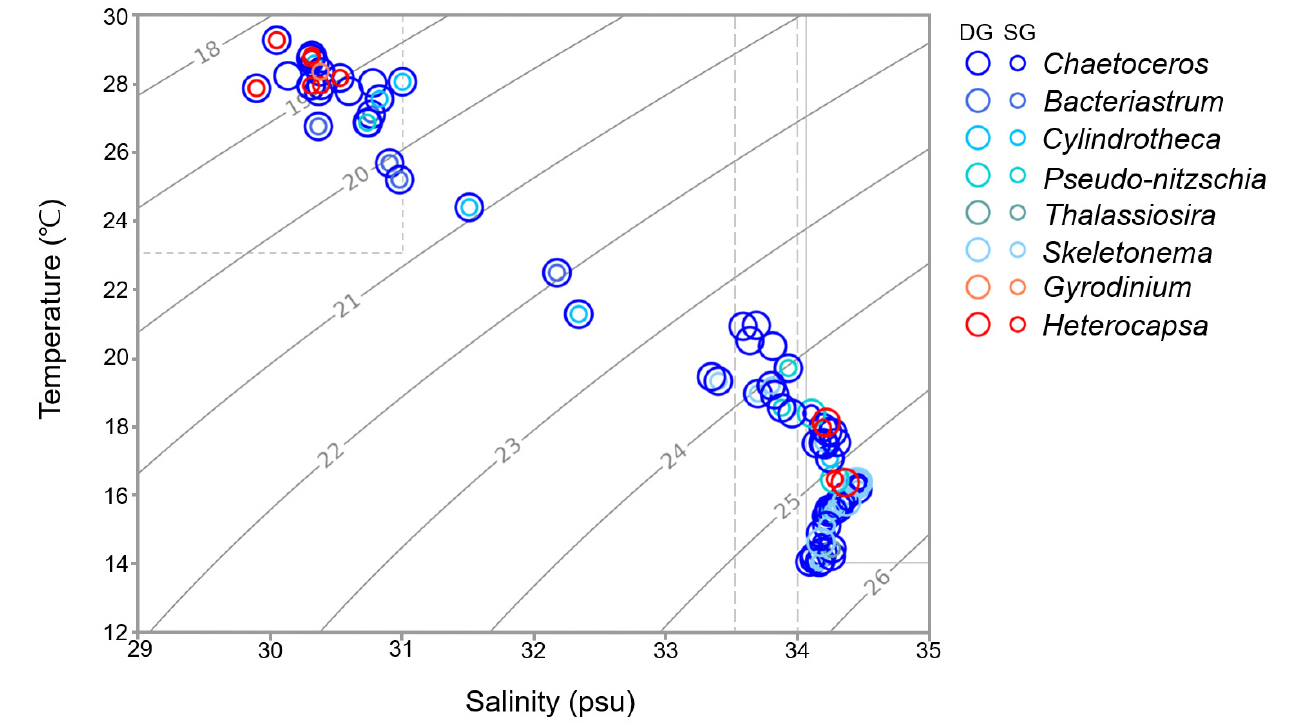
Fig. 8.
The dominant genus (DG) and subdominant genus (SG) points of each station are plotted on the Temperature - Salinity diagram. Water masses are defined by water temperature and salinity. Jeju Warm Current Water; above 12°C and 33.5–34 psu (gray dashed line), Tshshima Warm Current; above 14°C and above 34.1 psu (gray solid line) and Changiang River Diluted Water; above 23°C and below 31 psu (gray dotted line)
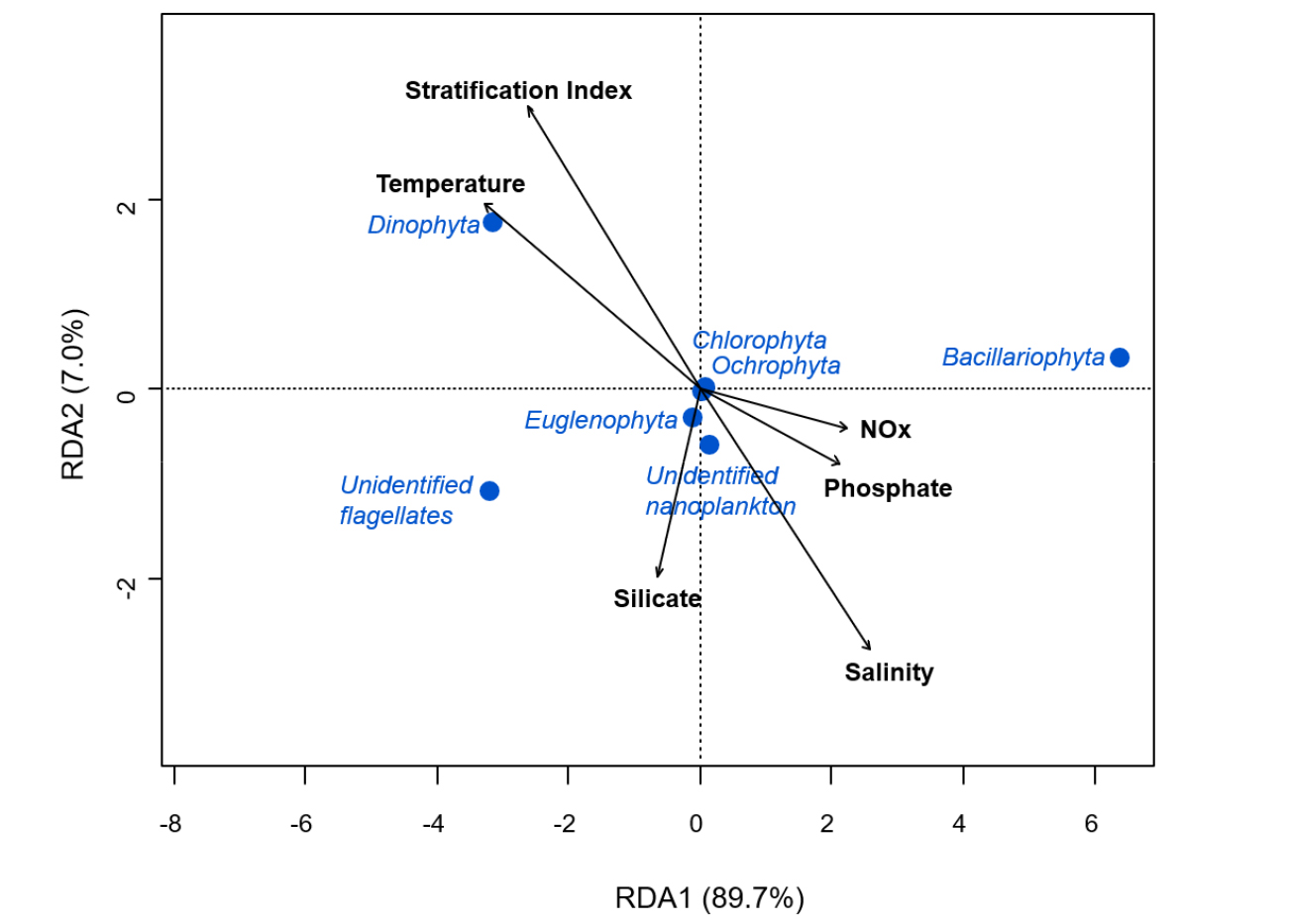
표층의 수온, 염분도, 영양염, SI 등 해양 환경요인과 식물플랑크톤 분류군별 출현과의 상관관계를 살펴보기 위해 RDA 분석을 실시하였다(Fig. 9). 분석 결과, 와편모조류(Dinophyta)의 출현은 수온 및 SI와의 강한 양의 상관성을 나타내었고, 염분 값과는 음의 상관 관계를 나타내었다. 규조류(Bacillariophyta)의 출현은 인산염 및 질산염+아질산염과 같은 식물플랑크톤 성장의 제한 인자로 작용하는 영양염들의 영향을 많이 받았음을 보여준다. 하지만 또 다른 규조류의 필수 영양염 중 하나인 규산염과는 상관관계가 없었다.
4. 토 의
2022 춘계에서 하계 제주 연안 표층수의 물리화학적 변동
조사기간 동안 춘계에서 하계로 갈수록 제주 연안의 평균 표층 수온은 높아지고 염분은 낮아지는 뚜렷한 경향을 보였다. 고 등 (2003)에 따르면 1998년과 1999년 제주 연안의 여름철 7월과 8월의 평균 수온 범위는 21–24.6°C로 보고되었으나, 본 연구에서의 월평균 수온은 26.8°C로 최소 2.2°C 이상 높게 측정되었다. 2015년 연구에서는 제주 연안의 8월 평균 수온은 25.6°C로 보고되었으며(박 등 2017), 본 연구에서의 8월 평균 수온은 27.6°C로 2°C 높게 측정되었다. 염분도의 경우 6월의 표층 평균 염분도는 34.2 psu에서 7월달 31.5 psu로 2.7 psu만큼 급격히 감소하였다. 제주도의 경우, 섬 전체가 투수성의 현무암층으로 이루어져 있어 여름철 많은 강우로 인한 연안 담지하수 유입이 큰 것으로 알려져 있다(황과 고 2012). 하지만 본 조사가 이루어지기 전 강우량의 양은 미비하였고(data not shown), 7월과 8월의 경우, 일부 정점을 제외하고 31–32 psu의 저염수가 수심 20 m까지 측정 되었기 때문에 강우로 인한 저염현상이 나타났다고 판단하기 어렵다. 7월의 경우, 제주 연안의 동쪽과 서쪽의 정점 간 수온 및 염분도 차이가 크게 측정되었는데, 이는 당시 제주 연안의 동쪽과 서쪽에 다른 수괴가 존재하였을 가능성을 시사한다. 위 사실들로 미루어 볼 때, 2022년 여름철 제주 연안은 고온・저염 특성의 양자강 희석수의 영향을 서쪽에서부터 강하게 받은 것으로 사료된다. Zhou et al. (2008)에 따르면, 1960년대부터 2000년대까지 중국의 급속한 경제 발전과 함께 양자강 배출수의 영양염 농도는 지속적으로 증가한 것으로 보고되었다. 이에 따라 중국 연안의 적조 발생 빈도가 급격히 증가하였는데, 적조의 대부분은 양자강 하구와 저장성(Zhejiang) 연안 지역에서 5월부터 6월까지 발생한 것으로 보고되었다. 하지만 본 연구의 영양염 분석 결과, 하계에 오히려 질산염+아질산염 및 인산염 등 주요 영양염의 평균 농도가 감소하였고, 7월 양자강의 영향을 강하게 받은 서쪽 정점과 그렇지 않은 동쪽 정점 간의 유의한 영양염 차이 역시 확인할 수 없었다. 김 등(2007)의 제주 남부 해역을 포함하는 동중국해 연구 결과에 따르면, 양자강과 인접한 서쪽 해역 정점에서 질산염+아질산염, 인산염 및 Chl.-a 농도가 높은 반면, 제주 인근 해역에서는 비교적 낮은 수치를 나타냈다. 이러한 사전 연구들과 본 연구의 영양염 분석결과를 토대로, 양자강 하구 인근의 양자강 희석수의 영양염 농도는 높지만, 제주도 해역으로 유입된 양자강 희석수에는 식물플랑크톤 성장에 필수적인 주요 영양염들이 이미 소비되거나 희석된 것으로 판단된다. 양자강 하구는 제주도와 약 450 km 가량 떨어져 있다.
또한 조사기간 동안 하계로 갈수록 인산염과 질산염+아질산염 농도는 하계로 갈수록 낮아지는 경향을 보였는데, 이는 4월과 5월 춘계 동안 번성한 식물플랑크톤의 소비에 의한 것으로 판단된다. 규산염의 경우 춘계의 규조류 대번성에도 불구하고 하계에도 춘계와 비슷한 농도를 유지하였다. 통계분석 결과 제주 연안의 동서남북 정점 간 각 영양염의 농도차는 유의하게 관측되지 않았다(data not shown). 큰 하천이 존재하지 않는 제주도의 지질학적 특성상 육상에서 하천으로 유입되는 영양염의 양은 제한적이며, 주요 영양염의 농도의 변동은 양식장 운영 등 인위적인 요인, 염/담 지하수의 유입량, 연안의 성층 형성, 주변 수괴의 특성 및 생물학적 요인 등에 의해 변동될 것으로 사료된다. 특히, 제주도는 투수성이 큰 현무암으로 구성되어 있어 강수량의 상당량은 지하수 및 용천수의 형태로 바다에 유출될 가능성이 높다. Kim et al. (2011)에 의하면 제주 화순만의 연안 용출수를 포함한 지하수 중 영양염(DIN, DIP, DSi) 평균 농도는 각각 120.9 µM, 1.22 µM, 97.6 µM, 방두만은 각각 71.6 µM, 2.1 µM, 259.2 µM로 측정되어 식물플랑크톤 일차 생산에 필요한 영양염(특히, 질산염+아질산염과 규산염) 공급에 큰 영향을 줄 것으로 판단된다. 방두만으로 공급되는 DIN, DIP, DSi의 플럭스를 계산해본 결과, 지하수에 의한 공급이 각각 90%, 20% 및 80%를 차지하는 것으로 조사되었다(Hwang et al. 2005). 특히, 규산염은 암석의 화학적 풍화작용과 가수분해에 의해 지하수로 공급되므로 제주도의 여름철과 같은 고온다습한 환경에서는 건조한 환경보다 지하수의 규산염의 함량이 높게 나타날 수 있다(박 2004). 제주도 해안에 분포되어 있는 해저 유출 지하수의 영양염 분석결과 일반 연안수에 비해 규산염의 농도가 약 5배 이상 높다고 보고하였다.
여름철 양자강 희석수 유입과 제주 연안 성층화에 미치는 영향
양자강 희석수는 여름철 강수량의 증가에 따라 양자강에서 유출된 많은 양의 담수가 연안의 해수와 뒤섞여 형성되며, 높은 수온과 낮은 염분도의 특성을 지닌다(Lie et al. 2003; Senjyu et al. 2006). 이러한 양자강 희석수는 통상 매년 6월경부터 남동풍의 영향을 받아 동쪽으로 확장되며, 약 1–2개월 후 제주해협을 통과하는 것으로 알려져 있다(김 등 1991; 김과 노 1994; Kim et al. 2009; Moon et al. 2009; 한국해양과학기술원 2016). 이로 인해 제주도 주변 해역에서는 고염분수인 대마난류수 영향을 받는 기간과 다르게 여름철에는 비교적 낮은 염분 농도가 관찰된다(Moon et al. 2009). 제주도 주변 해역으로의 양자강 희석수 유입은 대마난류수, 그 분지인 제주난류수 및 서해에서 남해로 연결되는 남해연안수 등의 해류에 영향을 받게 되는데, 이로 인해 양자강 희석수의 영향을 가장 빠르게 받는 지역은 제주도 서부이며 섬의 장벽 효과로 인해 해류의 흐름이 비교적 느린 제주도 북동부에서는 그 영향이 가장 약하고 느리게 나타난다(Moon et al. 2019; 차와 문 2020). 본 연구에서도 조사기간 중 7월부터 양자강 희석수의 영향이 나타났는데, 제주도 북동부 해역(J6, J7, J8)을 제외한 모든 정점의 표층에서 양자강 희석수의 특성(염분도 31 psu 이하 및 수온 23°C 이상)을 보이는 수괴가 관찰되었으며, 8월에는 모든 정점에서 관찰되었다. 제주도 북동부 해역은 위에서 언급한대로 해류 흐름이 비교적 약하며, 이 해역의 연안을 따라 형성된 강한 조류에 의해 수직혼합이 활발한 해역으로서(차와 문 2020), 이러한 요인이 복합적으로 작용하여 양자강 희석수의 영향이 상대적으로 더디게 나타난 것으로 판단된다.
일반적으로 성층 현상은 표층 수온의 상승이나 표층 염분도의 하강 등으로 인해 수괴 내의 연직 방향 밀도 변화 폭이 커질 때 강화된다. 조사기간 동안 양자강 희석수의 영향이 거의 없었던 6월의 경우, 제주도 주변해역의 수온 상승으로 인해 부분적으로 성층화가 진행되고 있었으나, 7월과 8월에는 아주 강한 성층화가 진행된 것으로 나타났다. 이는 계절 변화로 인한 제주도 해역의 자체적인 수온 상승의 원인도 있겠으나, 고온·저염의 양자강 수괴의 영향이 큰 것으로 판단된다. 이러한 결과는 양자강 기원의 저염분 수괴 유입이 북부 동중국해의 표층 수온을 증가시킨다는 연구 결과와도 일치한다(Moon et al. 2019).
한편, 강한 성층화가 진행된 7월과 8월에도 얕은 수심을 가진 정점(J1, J3, J4, J7)들에서는 낮은 SI 값이 측정되었다. 정점 J7의 경우 앞서 언급한 바와 같이 약한 해류 및 강한 조류에 의한 수직 혼합이 활발한 지역적 특성이 지배적인 것으로 판단되며, 다른 정점의 경우 비교적 수심이 얕은 지역으로서, 양자강 희석수의 영향을 표층부터 저층까지 전반적으로 받았기 때문으로 사료된다. 동중국해에서 관찰되는 양자강 희석수의 깊이는 표층부터 10–20 m 정도로 알려져 있기 때문에(Zhu et al 2022; Kim et al. 2023b), 수심이 얕은 지역은 양자강 희석수의 영향이 강하게 나타날 것으로 판단된다.
식물플랑크톤의 현존량 및 종조성 특징
일반적으로 춘계 온대해역에서는 표층수의 풍부한 영양염 및 일사량의 증가로 식물플랑크톤의 대번성이 발생하며, 하계에는 표층 수온의 증가로 성층이 발달하여 저층의 영양염 공급이 차단되어 대번성이 종료된다(Carstensen and Conley 2004). 2022년 제주 연안의 일차 생산량 변동은 전형적인 온대 해역의 특성을 잘 보여주었다. 평균 표층 수온이 15°C 이상으로 상승하기 시작한 4월과 5월에 높은 식물플랑크톤 현존량을 기록하였고, 성층이 발달하기 시작한 6월부터 현존량이 급감한 후 낮은 현존량이 8월까지 지속되었다. 조사기간 동안 식물플랑크톤의 현존량은 14,323–2,065,882 cells L-1의 범위로 측정되었는데, 2015년 조사 결과인 5,300–639,900 cells L-1 보다 3배 가까이 증가한 수치이지만, 1988년 제주 해안 조사에서 기록된 1,834–1,190,044 cells L-1의 현존량을 감안하였을 때(이 등 1990), 현존량 증감의 경향을 판단하기는 어렵다. 2022년 현존량 결과에서 특이한 점은 정점 간의 현존량 차이가 크다는 점인데 시기별로 정점 간 현존량은 적게는 약 9배에서 많게는 약 42배가량 차이를 보였다. 주로 현존량이 높은 정점들은 북쪽 및 동쪽에 존재하였다. 연안의 식물플랑크톤 현존량은 강우에 의한 육상 하천수 및 지하수의 유입, 심층수의 용승, 그리고 해류의 유입 등 다양한 경로를 통한 영양염류의 공급에 의존하게 된다(손과 백 2015; 권 등 2019). 앞서 언급한 바와 같이 제주 연안의 식물플랑크톤 현존량은 지하수로부터 공급되는 영양염류의 양에 의해 크게 영향을 받을 수 있는데, 박 (2004)의 연구에 따르면, 동쪽 지역의 해저 지하수의 해저 재순환량은 서쪽에 비해 1.25배 높은 것으로 산출되었고, 이에 따라 해저 지하수에 의해 유출되는 영양염의 농도 역시 동쪽 지역의 DIN은 약 1.71배, SiO2는 약 1.36배 서부 지역보다 높은 것으로 산출되었다. 또한 제주도에서 운영 중인 육상 양식장은 2019년 기준 424개로 주로 구좌읍과 성산읍, 표선면 등 동쪽 해안가에 밀집되어 있다(제주특별자치도 2021). 2020년 기준 양식장의 수는 지난 10년간 꾸준히 18% 증가하여(환경부 2023), 양식장 배출수는 제주 연안의 영양염 농도를 증가시키는 주요 원인 중 하나로 지적되고 있다. 양식장 배출수로 인한 제주 연안의 부영양화는 최근 제주 연안 갈파래류의 대번성에도 중요한 영향을 미치는 것으로 보고된 바 있다(이 등 2021).
Chl.-a 농도 분석 결과, 7월의 평균 Chl.-a 농도가 현미경으로 계측된 식물플랑크톤의 현존량보다 높게 측정되었는데, 이는 해당 시기 동쪽 지역 일부 정점(J6, J7, J9)에서 Chl.-a 농도가 3.73–4.28 µg L-1로 높게 측정되었다(Supplementary Fig. 1a). 식물플랑크톤 종의 크기 차이로 인해 정점 간 우점 종의 종류에 따라 식물플랑크톤의 현존량 대비 Chl.-a의 농도가 달라질 수 있으나, 해당 정점들의 우점 및 아우점 속은 다른 정점 들과 큰 차이점이 없었으며, 식물플랑크톤을 섭식하여 Chl.-a을 함유할 수 있는 Mesodinium 등의 동물성 플랑크톤들 역시 특별히 높은 개체수를 보이지 않았다(data not shown). 추가적으로 진행한 7월 동쪽과 서쪽 일부 정점을 대상으로 한 박테리아 군집 metagenomics 분석결과, 7월의 박테리아 군집은 대체적으로 남세균(cyanobacteria)이 우점하고 있었으며, 특히 동쪽 정점 J6과 J7의 경우, 남세균이 92.5%, 93.0%로 우점하고 있었다(Supplementary Fig. 1b). 우점종은 Cyanothece aeruginosa 였다. 제주 연안을 대상으로 한 박테리아 군집에 대한 사전연구는 많이 진행된 바 없으나, 2013년 1월, 5월, 9월, 12월 제주도 남부 지역에서 관측된 남세균의 최대 우점률은 3.59%에 그쳤다(Kim et al. 2023a). 본 연구에서는 박테리아의 현존량은 고려되지 않았으나, 현미경에서 계수되지 않았던 작은 남세균의 번성으로 인해 해당 시기 높은 Chl.-a 농도가 측정된 것으로 사료된다. 기후 변화로 인한 해수온 상승 등 해양 환경의 변화는 일차생산자인 식물플랑크톤의 군집 조성에 영향을 미치며, 환경 변화에 빠르게 대응하고 대사율이 높은 작은 크기의 식물플랑크톤이 점차 증가하는 경향이 있다는 사실이 알려졌다(Walther et al. 2002; Behrenfeld et al. 2006; 손 등 2012; 장 등 2006). 따라서, 향후 아열대 해양환경으로 변화 중인 제주 연안의 일차생산 조사는 이러한 작은 크기의 일차생산자인 남세균의 현존량 및 종조성 또한 반드시 고려되어야 할 것으로 판단된다.
조사 기간 동안 제주 연안 해역은 Chaetoceros, Skeletonema 속 등의 규조류가 평균 70.1%로 우점하였다. 2015년 연구에서도 제주 연안에서의 규조류 점유율은 51–91% 범위로 나타난 것으로 조사되어 규산염이 풍부한 제주 연안에서 규조류는 일차생산을 담당하는 주요 분류군인 것으로 판단된다(김 등 2019). 상대적으로 규조류는 다른 식물플랑크톤 분류군에 비해 성장 속도가 빨라 영양염의 제한이 없는 환경에서 우점하여 출현할 수 있다(Egge and Aksnes 1992). RDA 분석 결과, 조사 기간 동안 규조류의 우점현상은 규산염 농도와는 무관한 것으로 나타났는데, 조사 기간 동안 제주 연안에서 풍부한 농도의 규산염이 규조류 성장의 제한요인으로 작용하지 않는 것으로 판단되며, 이는 제주에서 규조류가 항상 높은 우점률을 보일 수 있는 이유로 사료된다.
조사 기간 동안 와편모조류와 편모조류의 점유율이 하계로 갈수록 증가하는 경향이 있는 것을 확인할 수 있었다. RDA 분석 결과, 기타 편모조류의 경우 측정한 해양 물리화학적 환경요인과 별다른 출현 특성을 확인할 수는 없었지만, 와편모조류 출현의 경우 성장에 필요한 영양염이 아닌 수온 및 성층화 정도와 연관성이 큰 것을 확인하였다. 편모조류의 출현비율이 상대적으로 높았던 7월과 8월 여름철 표층의 영양염류의 농도는 낮았다. 빈영양 수괴에서 와편모조류는 편모를 활용한 일주 수직 운동을 통해 상대적으로 풍부한 저층의 영양염을 소비할 수 있기 때문에 성층화가 발달된 해역에서 규조류를 포함하는 단순 부유성 미세조류들보다 개체수를 증가시킬 수 있는 유리한 생태적 전략을 가진다(Jeong et al. 2015; Lim et al. 2022). 2022년 여름철은 양자강 희석수에 의해 성층화가 강화되었고, T-S 다이어그램 분석 결과, 양자강 희석수의 영향을 받은 시기 및 지역에서 와편모조류인 Heterocapsa의 출현이 증가한 것을 확인할 수 있었다. 8월의 경우 Heterocapsa 및 Gyrodinium 이 동쪽의 정점(J7–J9)를 제외하고 모든 정점에서 아우점속으로 출현하였다. 해당 생물들의 출현은 해류 및 수괴의 이동 등 해양 물리적 요인들이 복합적으로 영향을 주고 있는 것으로 사료된다. 2009–2011년 대마난류 등의 해류의 영향에 의해 Cochlodinium, Pseudo- nitzschia 등 외래 유해 적조원인 생물 종의 개체군이 유입하고 있다는 보고가 있었으며(Kim et al. 2015, 2018; Park et al. 2014), 특히, 2016년과 2017년 여름철인 8월 남해안 조사결과에서도 양자강 희석수의 영향을 받아 연안 수괴의 성층 현상이 강화되었고, 2016년에는 Gymnodinium, Gyrodinium, Prorocentrum 속이 2017년에는 Alexandrium, Cochlodinium 속 등의 와편모조류가 우점하는 현상이 관측되어 양자강 희석수가 유해 적조 생물의 출현 결정에 주요한 영향을 미칠 수 있다고 보고되었다(Kim et al. 2019; Lim et al. 2019).
또한, 4월을 보면 서쪽에서 북쪽 정점(J2–J5)에서는 Chaetoceros가 평균 61% 이상 우점, Skeletonema가 18.7% 정도로 아우점 속으로 나타났으나 동쪽에서 남쪽 정점(J8–J12)에서는 반대로 Chaetoceros의 점유율이 평균 14.8%로 줄었고 Skeletonema가 35.4%로 증가했다. 7월에도 전 정점에서 주로 Chaetoceros 속이 우점하였으나, 서쪽에서 북쪽 정점들의 경우 Bacteriastrum 속이 아우점 속으로 출현했던 반면, 동쪽에서 남쪽 정점들에서는 Cylindrotheca가 아우점 속으로 주로 출현하였다. 이렇듯 식물플랑크톤의 출현 양상이 서쪽에서 북쪽, 그리고 동쪽에서 남쪽 정점들끼리 해역별로 유사한 사실은 2015년 사전연구에서도 이미 보고된 바 있으며 이는 제주 연안에 미치는 대마난류, 제주난류, 양자강 유출수의 복합적인 계절적 영향으로 유추하였다(김 등 2019).
5. 요약 및 결론
2022년 춘계에서 하계 제주 연안 식물플랑크톤 군집은 대체적으로 규조류가 우점하였으나, 하계로 갈수록 고온・저염의 양자강 희석수의 영향을 강하게 받으며 성층 현상이 강화되었고, 이로 인한 표층수 영양염의 고갈현상과 함께 규조류의 성장은 둔화되고 상대적으로 저층의 영양염 활용이 가능한 편모조류의 비율이 증가한 것으로 보인다. 제주도의 지질학적 특성상 연안에 규산염이 풍부하기 때문에, 규조류가 제주 연안의 주요 일차생산자 역할을 수행하는 것으로 예상된다. 또한 해역 별로 보면 제주도의 서쪽과 북쪽, 그리고 동쪽과 남쪽 해역의 식물플랑크톤 군집이 유사한 경향을 나타냈다. 이렇듯 시공간에 따라 제주 연안 식물플랑크톤의 출현 특성이 나타나는 이유는 제주 연안이 계절에 따라 대마난류, 제주난류 및 양자강 희석수 등 해양 물리화학적 특성이 서로 다른 다양한 수괴들의 영향을 복합적으로 받기 때문으로 보인다. 본 연구를 통해 최근 춘계와 하계 제주 연안의 식물플랑크톤 현존량 및 종 다양성 정보를 업데이트 하였으며, 계절에 따른 다양한 수괴, 특히 대마난류와 양자강 희석수의 영향으로 인한 식물플랑크톤 군집의 변동 가능성을 확인하였다(Supplementary Fig. 2). 본 연구는 춘계에서 하계까지의 조사만 수행했으므로 향후 추계와 동계의 조사 및 분석 또한 함께 고려되어야 할 것으로 보이며, 작은 크기의 식물플랑크톤들 특히 남세균에 의한 높은 일차생산이 예측되는 바, 제주 연안 생태계의 보다 나은 이해를 위해 이들을 포함하는 면밀한 조사 체계가 필요하다.
