Copyright © 2023 Ocean and Polar Research
ABSTRACT
1. 서 론
지난 제4기 동안 지구의 환경은 빙하기–간빙기에 따라 변화를 반복하였고, 이러한 변화는 북반구 고위도 지역의 여름철 동안 태양 복사에너지의 흡수를 조절하는 궤도 강제력에 의해 조절된다(Berger 1988). 태양 복사에너지의 변화는 밀란코비치 기작(Milankovitch mechanism)에 의해 설명되고, 지구 궤도 변수인 세차운동(precession), 축 경사(obliquity), 이심률(eccentricity)의 규칙적인 변화에 의해 각각 2만 년, 4만 년, 10만 년 주기로 간빙기와 빙하기가 반복된다(Paillard 2001). 지구의 환경은 간빙기와 빙하기에 따라 급격하게 변화하며 두 시기는 매우 다른 특징을 보여준다. 빙하기 동안 북유럽과 북미의 대륙에는 두꺼운 빙상이 덮여 있었으며, 이러한 대륙빙상의 형성은 해수면의 하강을 야기하였다(Batchelor et al. 2019). 또한, 빙상은 알베도를 증가시켜 태양으로부터 유입되는 복사에너지를 더 많이 반사하기 때문에 대기와 해수의 온도가 하강하였다(Siddall et al. 2003; Pahnke and Sachs 2006). 최근의 모델 연구들은 빙하기–간빙기 동안 대륙빙상의 규모, 해빙의 확장과 감소, 표층수의 성층화, 바람의 세기와 방향 변화 등의 복잡한 상호작용에 의해서 대기 이산화탄소 농도의 변동이 있었음을 밝혔다(Kohfeld et al. 2005; Köhler et al. 2005; Martínez-Garcia et al. 2009). 남극대륙의 4개 지역에서 시추한 빙하 코어(Vostok, EPICA Dome C, EPICA Dronning Maud Land, Talos Dome)를 사용하여 과거 기후 이벤트 복원 연구에서 일관된 척도로 사용되는 AICC (Antarctic Ice Core Chronology)에 의하면 지난 마지막 빙하기 동안 대기 이산화탄소 농도는 약 180 ppm으로 간빙기보다 80에서 100 ppm 정도 낮았다고 보고되었다(Petit et al. 1999; Veres et al. 2013). 빙하기–간빙기의 대기 이산화탄소 농도 차이를 설명할 수 있는 기작은 아직 완전히 밝혀지지 않았으나, 빙하기 동안 남극해의 표층 해양에서 일차생산의 변화는 대기 이산화탄소 농도 변화에 잠재적으로 중요한 역할을 했다고 알려진 바 있다(Martin 1990; Moore et al. 2000; Petrou et al. 2016). 표층 해양의 일차생산성 증가와 그로 인한 내보내기 생산(export producion)의 증가는 표층 해양의 탄소를 심해로 제거하여 대기 이산화탄소의 농도를 감소시킨다(Kumar et al. 1995; Marinov et al. 2006).
표층 해양의 고생산성 복원을 위해 미화석 군집 분석과 같은 미고생물학 연구와 동위원소 또는 미량 금속의 지화학적 측정을 포함한 다양한 프록시가 사용되고 있다(Lazarus and Caulet 1993; Latimer 2004). 그중에서도 해양 퇴적물에서 분석된 생물기원 오팔 함량은 과거 표층 생산성의 변화를 복원하는 데 광범위하게 이용된다. 생물기원 오팔 퇴적의 기록들은 지난 15 Myr 동안 다양한 해양에서 오팔 퇴적에 대한 기후, 지구조적, 해양학적, 생물학적 사건 사이의 상호작용을 포함하여 퇴적 기작, 과정, 결과 그리고 양상의 변화에 대한 정보를 제공한다(Ragueneau et al. 2000; Cortese et al. 2004). Ragueneau et al. (2000)은 일반적으로 대서양보다 태평양 분지에서 퇴적물 내 오팔 보존의 상태가 양호하며, 오팔의 퇴적은 다른 해역보다 남극해 주변에서 우세하다고 보고하였다. 오팔로 구성된 각을 형성하는 일차생산자인 규조류는 해양 생태계에서 일차생산의 약 40%를 담당한다(Treguer et al. 1995). 표층 해양에서 규조류의 생산성은 심해로 유기탄소를 운반하고, 다양한 시간 규모에서 대기 중 이산화탄소 농도의 변화와 관계된다(Cortese et al. 2004). 남극해는 고생산성의 변화에 따라 규조류에 의한 이산화탄소의 생물학적 펌프 작용이 활발하며. 이로 인해 대기–해양 이산화탄소 교환에서 중요성을 갖는다(Anderson et al. 2009).
남극해는 형성 이후 전지구 해양 순환, 열 수송, 생물학적 생산성 그리고 대기와의 이산화탄소 교환에 중요한 역할을 하고 있으며, 특히 남극해의 생지화학적 시스템에서 일차생산자에 의한 탄소 순환은 표층 해수의 영향을 받는다(Latimer 2004; Marinov et al. 2006; Gruber et al. 2009; Henley et al. 2020). 따라서 과거 기후 변화에 대한 남극해 표층 해수의 특성 변화와 그 역할을 이해하는 것은 필수적이다. 이러한 중요성에도 불구하고 남극해의 인도양 해역에서 고생산성 연구는 아직까지 많이 부족한 실정이다(Manoj and Thamban 2015). 이 연구는 마지막 빙하기부터 홀로세까지 남극해 인도양 해역의 생물기원 오팔 함량 변화를 통한 표층 생산성의 변화를 이해하고 남극해 인도양 해역의 고해양 환경 변화를 복원하는 것을 목적으로 한다.
2. 재료 및 방법
연구 지역
전선(front)은 해수 특성의 차이가 뚜렷한 두 수괴들의 경계로 정의된다(Gordon 1971). 남극해에는 아열대 전선(Subtropical Front), 아남극 전선(Subantarctic Front), 남극 전선(Antarctic Polar Front) 그리고 남극순환류의 남쪽 경계 전선(Southern ACC Front)이 주요 전선을 형성한다(Fig. 1; Whitworth and Nowlin 1987; Orsi et al. 1995). 아열대 전선은 아열대 표층수와 아남극 표층수 사이의 경계로 –40°S에 위치한다. 아남극 전선의 위치는 남극중층수(Antarctic Intermediate Water)가 하강하는 지역이며 남극순환류의 북쪽 경계로도 정의된다. Belkin and Gordon (1996)에 의해 남극 전선은 아표층 해수의 최소 온도가 2°C인 최북단 영역으로 정의되었다. 남극 해역(Antarctic Zone)은 남극 전선과 남극순환류의 남쪽 경계 전선 사이에 위치하는 영역으로 이 해역의 성층화는 온도보다 염분에 의해 제어된다(Pollard et al. 2002). 연구지역인 콘래드 해령에서 얻은 코어 COR-1bPC의 위치는 남극 전선의 남쪽에서 채취되어 남극 해역에 위치한다(Fig. 1).
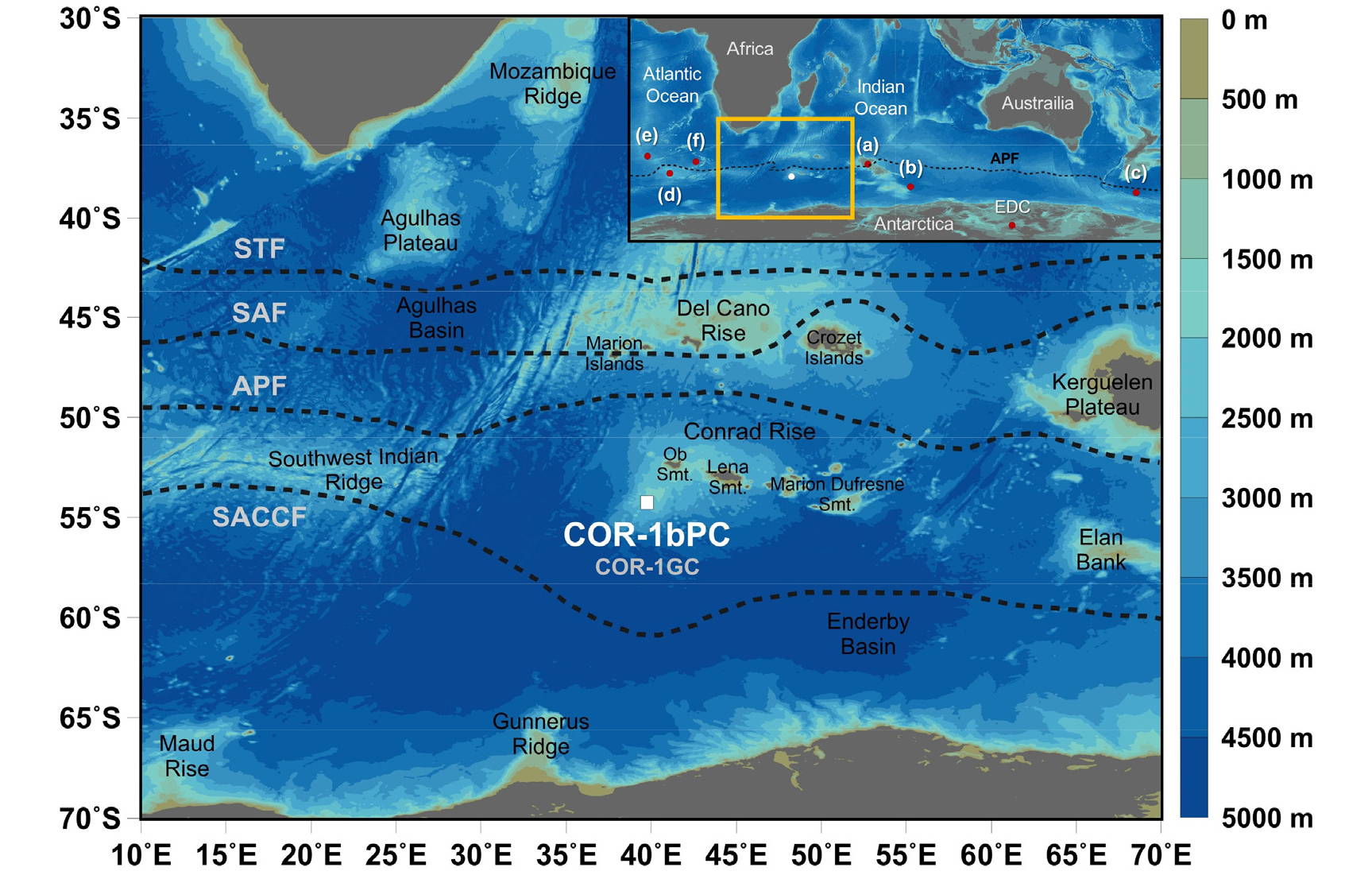
Fig. 1.
코어 기본 정보
이 연구에 사용된 코어 COR-1bPC (길이 10.47 m)는 남극해의 인도양 해역에 위치한 콘래드 해령 남서쪽 사면(54°16.04’S, 39°45.98’E; 수심 2,828 m)에서 R/V Hakuho-maru호를 이용하여 2010년 12월에 수행된 KH-10-07 탐사에서 획득되었다(Fig. 1). 퇴적물 표면에서 코어 퇴적물의 밝기를 의미하는 L* 값의 측정을 위해 Minolta사의 CM-2002 photospectrometer를 사용하여 1 cm 간격으로 측정하였다. L* 값은 Oiwane et al. (2014)에서 발표되었다. 코어 퇴적물의 대자율(Magnetic susceptibility) 값은 multi-sensor core logger (MSCL-S, GEOTEK Ltd.; sensor, Bartington Instrument Ltd.)를 이용하여 2 cm 간격으로 측정하였다. 분석 실험에 사용되는 부시료는 채취 후 동결건조기를 이용하여 건조하였으며, 볼밀을 사용하여 분말화하였다.
생물기원 오팔 함량 측정
퇴적물의 생물기원 오팔 함량은 약 10 cm 간격으로 채취된 113개의 부시료에서 분석되었다. 퇴적물의 생규소(biogenic silica, B-Si) 함량을 측정하기 위해 Mortlock and Froelich (1989)와 DeMaster (1981)의 방법을 변형시킨 습식 알카라인 연속 추출법(wet alkaline sequential extraction)을 사용하였다. 50 ml 코니칼 튜브에 분말화된 시료 약 15 mg을 넣고 1N Na2CO3를 30 ml 씩 주입한 후, 85°C로 설정된 항온 수조기에 넣는다. 그 후 2, 3, 4, 5시간마다 튜브 속의 용액을 100 μl씩 추출하여 2 ml의 0.1N HCl이 들어있는 바이얼에 넣는 과정을 총 4번 반복한다. 추출된 용액이 들어있는 바이얼에 15분 간격으로 ammonium molybdate solution과 reducing solution을 넣고 12시간 동안 안정화를 시킨다. 그 후 분광분석기(Biochrom Libra S22 Spectrophotometer)를 사용하여 812 nm 파장에서 생규소의 함량을 측정하였다. 측정된 생규소 함량에 규소와 오팔의 비인 2.4를 곱해주어 생물기원 오팔 함량을 계산하였다.
Biogenic opal (%) = B-Si (%) × 2.4
연대 설정 및 퇴적률
코어 COR-1bPC의 퇴적 연대는 Oiwane et al. (2014)에 의해 발표되었다. 4개의 방사성 탄소 연대는 부유성 유공충(Neogloboquadrina pachyderma sinistral)을 이용하여 가속기 질량 분석(Accelerator Mass Spectrometry) 14C 연대 측정법에 의해 도쿄대학교에서 획득되었다(Fig. 2). 측정된 탄소 연대는 남극해의 저장 효과(890 yr)를 고려하여(Bard 1988), 보정 프로그램인 CALIB 6.1.0을 사용하여 역년(cal yr BP)으로 변환되었다(Stuiver and Reimer 1993). 또한, EPICA Dome C (EDC) 빙하 코어의 온도 자료와 L* 사이의 비교를 통해 추가적인 연대를 설정하였다(Fig. 2). 코어 COR-1bPC의 깊이 4.7 m에서 L* 값의 급감은 퇴빙기의 시작을 나타내는 EDC 빙하 코어의 17.4 ka에 해당하는 깊이와 대비된다. 이러한 탄소 연대 측정과 층서 대비에 의해 코어 COR-1bPC의 최하부는 약 43,000 cal yr BP로 예상된다.
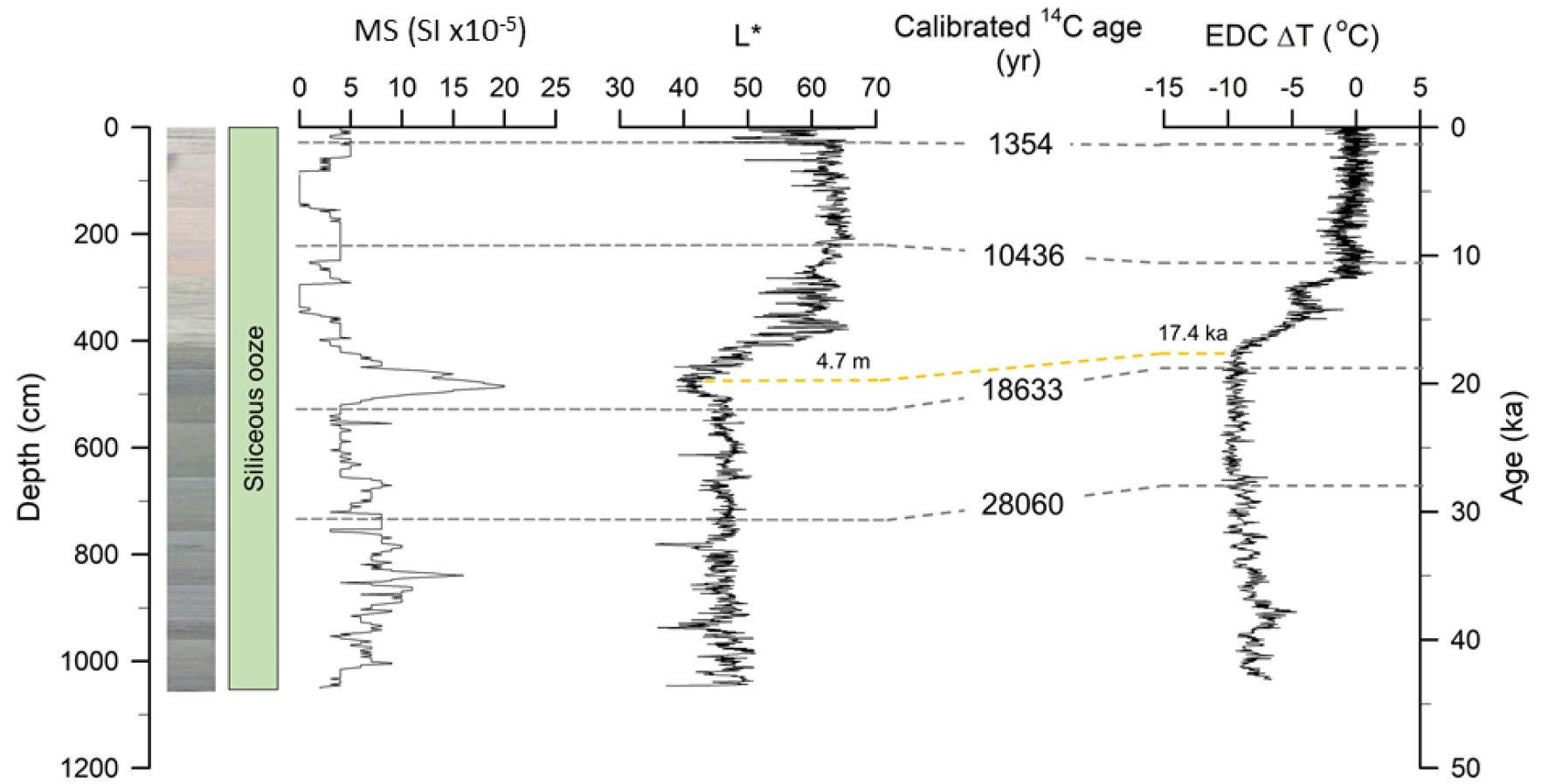
Fig. 2.
Downcore variation of magnetic susceptibility (MS), lightness (L*) of COR-1bPC with the stratigraphic photograph and sediment column. Four AMS 14C age data and L* were reported by Oiwane et al. (2014). An additional age at 4.7 m was estimated by the correlation between L* and EPICA Dome C (EDC) ΔT (°C) (Jouzel et al. 2007)
3. 연구결과
콘래드 해령에서 채취된 코어 COR-1bPC의 퇴적물은 생물기원 규조의 함량이 30% 이상인 규조 연니로 분류된다(Fig. 2). 퇴적물의 L*값은 평균 52.0%으로, 코어 최하부에서 4.7 m까지 평균 46.0%의 어두운 값을 갖다가 4.7 m에서 코어 최상부까지 59.4%으로 증가하며 점차 밝아진다. 퇴적물에 포함된 자성 물질의 양을 의미하는 대자율 값의 평균은 5.28(10-5 SI)이다. 급격한 변화를 보이는 4.7 m 기준으로 코어를 상부와 하부로 나누었을 때, 상부의 대자율 평균은 3.26(10-5 SI), 하부의 대자율 평균은 6.91(10-5 SI)이다. 어두운색을 보이며 L* 값(40–50)이 낮은 코어 하부의 대자율은 약 10(10-5 SI) 내외로 변화하고, 밝은색을 보이며 L* 값이 높은(> 60) 코어 상부의 대자율은 5(10-5 SI) 이하로 나타난다. 남극해에서 획득된 다른 퇴적물 코어의 대자율과 비교했을 때, 이는 매우 낮은 값에 속한다(e.g. Crosta et al. 2002; Thamban et al. 2005; Nielsen et al. 2007). 전반적으로 코어 퇴적물의 대자율 값이 15(10-5 SI) 이하로 작게 나타나는 것은 생물기원 입자가 우세하게 구성되고, 자성을 가지는 쇄설성 입자의 함량이 적기 때문이다(Fig. 2). 한편 코어 하부의 높은 대자율은 자성을 보이는 쇄설성 입자의 함량이 상대적으로 많기 때문으로 해석된다. 코어 중앙부에 속하는 4.7 m 지점의 퇴적물의 대자율은 최대값, L* 값은 최소값에 해당한다는 특징을 보인다. Oiwane et al. (2014)가 발표한 전밀도(bulk density)는 코어 최하부인 10.5 m에서 5.0 m까지 1.2–1.3 g/cm3이고, 5.0 m에서 2.5 m 사이에서 1.1 g/cm3로 변하며, 2.5 m에서 해저면인 코어 최상부까지는 1.0–1.1 g/cm3으로 비교적 작은 값을 보이며 전체적으로 코어 상부로 가면서 감소한다. 따라서 퇴적물의 대자율, L* 값, 그리고 전밀도를 포함하는 퇴적물의 특징은 코어 하부에서 상부까지 거의 동일한 변화를 보여주며 코어 중앙에 큰 변화가 특징적으로 나타난다(Fig. 2).
Oiwane et al. (2014)의 연대 자료를 이용하여 계산한 COR-1bPC의 선형 퇴적률은 코어 상부(43–18 cal kyr BP)와 하부(11–0 cal kyr BP)에서 20–23 cm/kyr로 비슷하며, 코어의 중앙부(18–11 cal kyr BP)는 39 cm/kyr로 약 1.5에서 2배 정도 높다(Fig. 3). 코어 COR-1bPC 상부의 퇴적률은 코어 채취 지역 인근에서 발표된 코어 COR-1GC (54.27˚S, 39.77˚E; 수심 2,834 m; Fig. 1)의 퇴적률(19 cm/kyr)과 매우 유사하다(Fig. 3; Orme et al. 2020). 일반적인 원양성 퇴적물의 퇴적률은 1 cm/kyr 정도로 매우 낮다(Pickering and Hiscott 2015; Rotzien et al. 2022). 콘래드 해령의 퇴적률이 일반적인 원양성 퇴적물의 경우보다 높은 것은 남극순환류에서 영양염의 활발한 용승으로 인해 규조류가 높은 생산성을 보이는 것과 콘래드 해령 서쪽에서 남극순환류의 사행으로 인한 퇴적물의 이동 때문으로 제안되었다(Durgadoo et al. 2008).
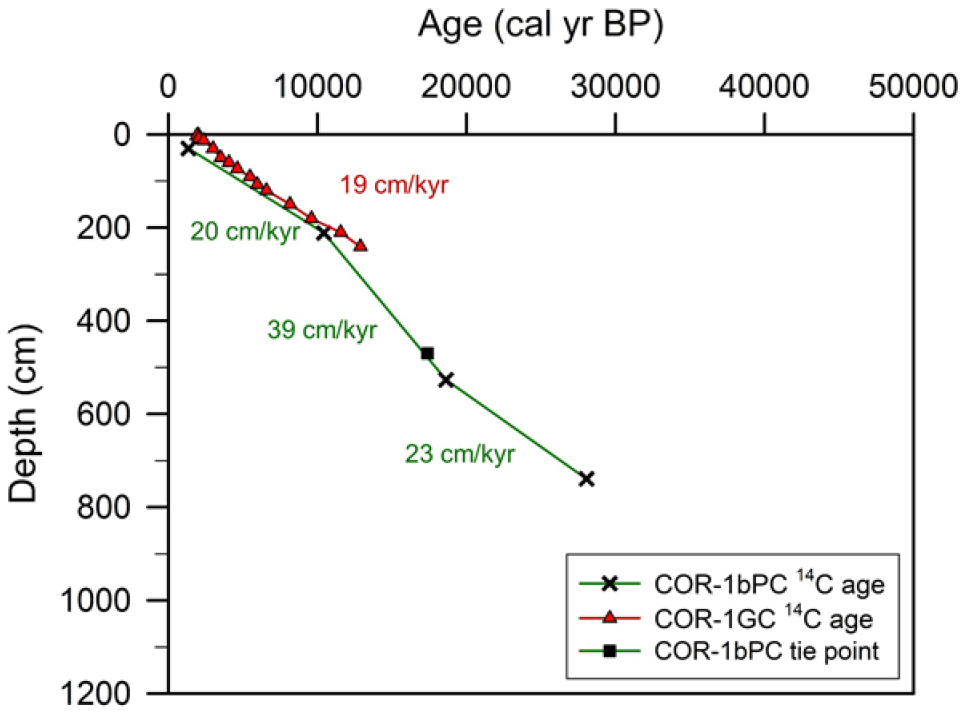
Fig. 3.
Sedimentation rates (green line) of core COR-1bPC are divided into the three (upper, middle, and lower) parts. The sedimentation rate in the upper part of core COR-1bPC is similar to that (red line) of COR-1GC collected in the same area (Orme et al. 2020)
코어 COR-1bPC 퇴적물의 생물기원 오팔 함량은 코어의 하부에서 상부로 가면서 전반적으로 증가한다(Fig. 4). 생물기원 오팔 함량은 최소 20.4%와 최대 85.4% 사이에서 변화하며, 생물기원 오팔 함량의 최솟값이 나타나는 시기는 마지막 빙하기로 20.4%이고, 최댓값이 나타나는 시기는 홀로세에 해당하며 85.4%이다. 전 구간인 약 43,000년 동안 오팔 함량의 평균은 50.8%로 대부분의 퇴적물이 규질 연니로 이루어졌음을 지시한다. 약 16,500년에서 13,500년 사이에 생물기원 오팔 함량은 30.6%에서 82.0%로 급격한 증가를 보인다. 이 구간은 퇴적물의 다른 특성(대자율, L*, 전밀도)의 변화 구간과 일치한다(Fig. 2). 코어 COR-1bPC 퇴적물의 생물기원 오팔 함량은 지난 43,000년 동안 EDC ΔT와 AICC (Antarctic Ice Core Chronology) CO2 농도와 유사한 변화를 보인다(Fig. 4; Veres et al. 2013; Bereiter et al. 2015). 생물기원 오팔의 평균 함량은 마지막 빙하기(43–18 cal kyr BP) 동안 38.9%이며 퇴빙기(18–11 cal kyr BP) 동안 58.2%로 증가하였다. EDC ΔT와 AICC CO2 농도가 높은 홀로세(11–0 cal kyr BP) 동안 생물기원 오팔의 평균 함량은 73.4%으로 마지막 빙하기 이후 거의 두 배 정도 증가하였고 퇴빙기에 급격한 증가를 보였다. 오팔 함량의 변동폭은 퇴빙기 동안 가장 크며, 퇴빙기 후반에 나타나는 오팔 함량의 급격한 증가는 EDC ΔT와 AICC CO2 농도의 증가와도 일치한다(Fig. 4).
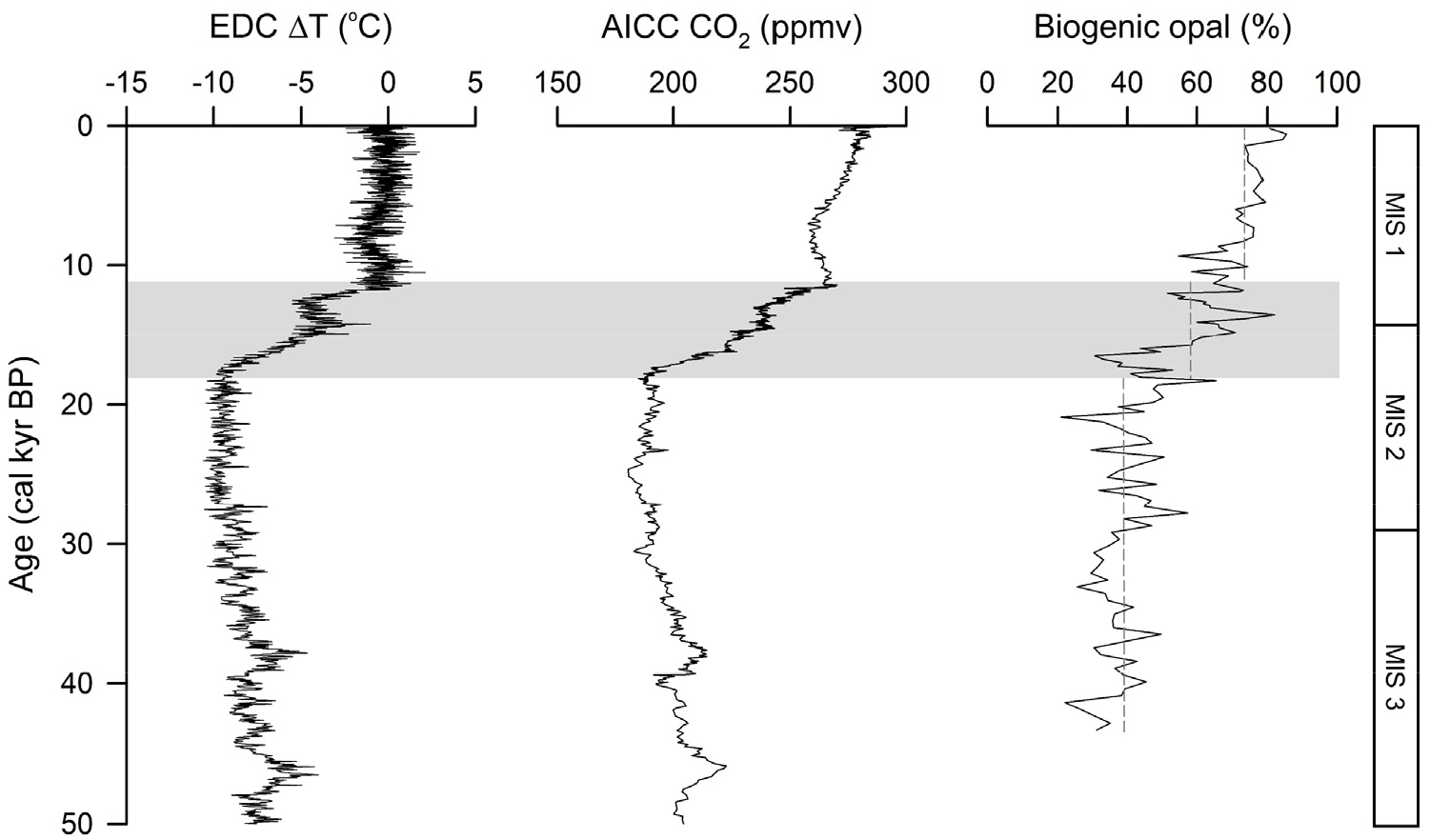
Fig. 4.
Downcore profile of EPICA Dome C (EDC) ΔT (°C) (Jouzel et al. 2007), Antarctic Ice Core Chronology (AICC) CO2 concentration (Petit et al. 1999; Veres et al. 2013), and biogenic opal content of core COR-1bPC. The dotted gray vertical lines represent the average of the biogenic opal content during the last glacial period, deglaciation, and Holocene
4. 토 의
남극 전선 주변해역의 빙하기 이후 규조 생산성의 변화
남극해 남극 전선의 북쪽에 위치한 아남극 해역(Subantarctic Zone)과 남극 전선 남쪽에 위치한 남극 해역에서 지난 마지막 빙하기 동안 표층 해수의 일차 생산성 변화는 반대의 양상으로 나타난다. 아남극 해역에서는 빙하기 동안 일차생산이 증가하는 반면, 남극 해역에서는 감소하며 간빙기에는 반대의 양상이 나타난다(e.g. Charles et al. 1991; Francois et al. 1997; Anderson et al. 2009; Bradtmiller et al. 2009). 남극 해역에서 일차 생산성의 최대 ~75%까지 기여하는 규조류는 주로 겨울철 해빙의 북쪽 경계와 남극 전선 사이에 퇴적되어 규조 연니대(diatom ooze belt)를 형성한다(Burckle and Cirilli 1987; Crosta et al. 2005). 일반적으로 오늘날 남극 전선의 남쪽에 위치한 규조 연니대에는 규조류의 퇴적이 우세하지만, 빙하기 동안 급격하게 감소하는 특징이 나타난다(e.g. Cooke and Hays 1982; Kumar et al. 1993, 1995; Bareille et al. 1998). 남극 전선의 남쪽에 위치한 콘래드 해령에서 채취한 코어 COR-1bPC의 생물기원 오팔 함량 또한 빙하기에 낮은 값을 보이다가 홀로세에 증가하는 양상을 보인다(Fig. 5). 이러한 빙하기–간빙기 오팔 함량 변화는 이전 연구결과들과 일치하며, 빙하기–간빙기 기후 변동성을 특성화한 EDC ΔT의 변화와도 유사하다(Fig. 4).
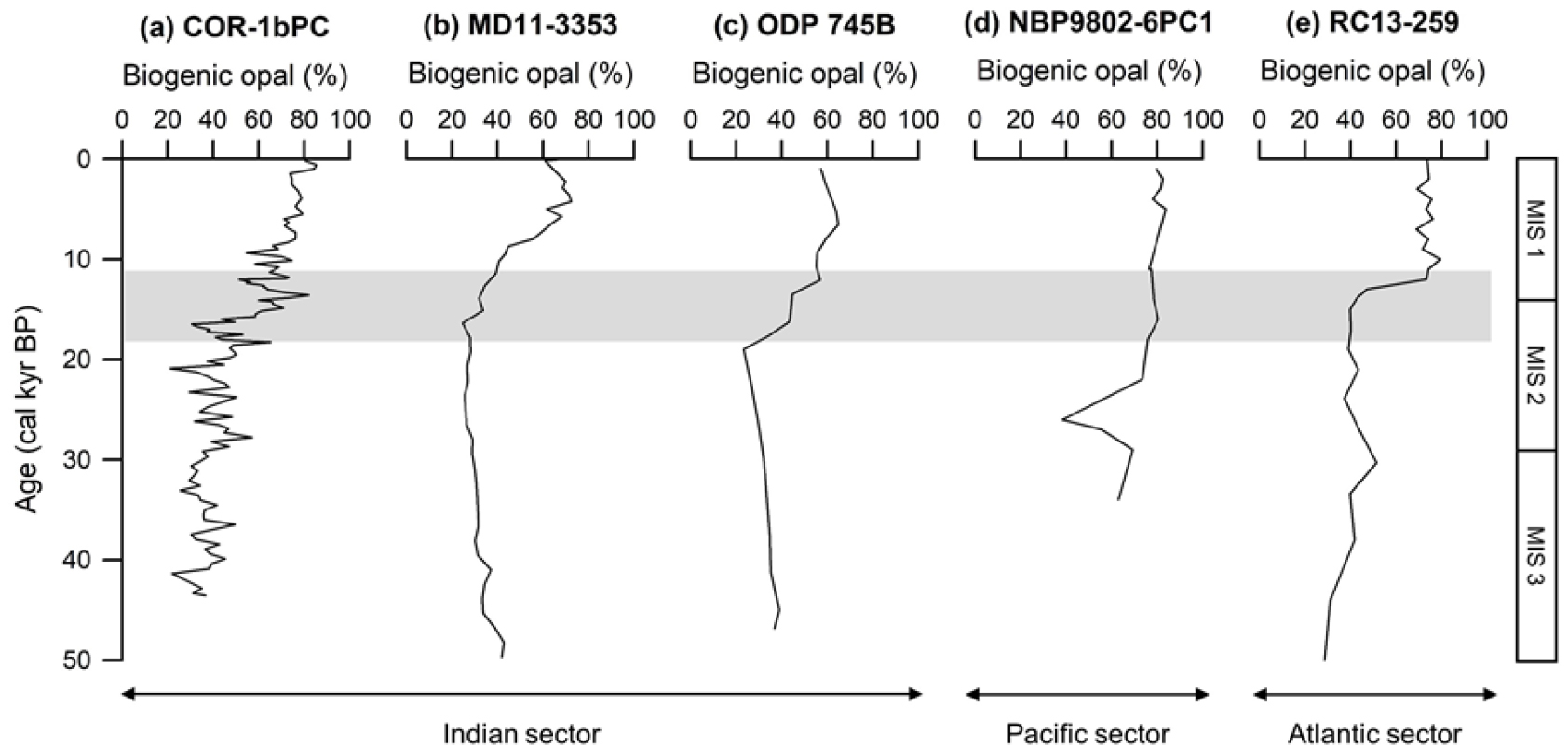
Fig. 5.
연구지역인 콘래드 해령에서 채취한 코어 COR-1bPC처럼 남극 전선의 남쪽에서 획득된 코어들의 생물기원 오팔 함량은 서로 유사한 결과를 보인다(Fig. 5). 콘래드 해령의 동쪽에 위치한 남인도양 해역의 케르굴렌 고원에서 획득된 코어 MD11-3353(50.57˚S, 68.39˚E; 수심 1,568 m)는 현재 남극전선의 북쪽에 위치하지만, 전선과의 거리가 매우 가까워 남극 전선 남쪽과 유사한 생물기원 오팔 함량 변화를 보인다. 마지막 빙하기부터 퇴빙기까지 생물기원 오팔 함량의 평균값은 31.9%이며, 홀로세 동안의 평균값은 61.0%으로 증가한다(Civel-Mazens et al. 2021; Fig. 5a). 마찬가지로 남인도양의 케르굴렌 고원과 호주–남극 분지(Australian–Antarctic Basin)의 경계에서 시추된 ODP Site 745B (59.62˚S, 86.87˚E; 수심 4,082 m)는 생물기원 오팔 함량의 평균값이 마지막 빙하기 동안 32.1%이며, 퇴빙기 이후 53.2%로 증가한다(Fig. 5b; Kaiser et al. 2021). 남태평양의 남극 전선 남쪽에 위치한 캠벨 고원(Campbell Plateau) 남동쪽에서 획득한 코어 NBP9802-6PC1(61.88˚S, 169.98˚W; 수심 3,245 m)도 마지막 최대 빙하기(Last Glacial Maximum) 동안 생물기원 오팔 함량이 급격하게 감소한 이후 점차 증가하는 양상을 보인다(Fig. 5c; Chase et al. 2003). 마지막으로 남대서양에 위치한 남극 전선 남쪽의 남미–남극 해령(South America–Antarctic Ridge)에서 획득된 코어 RC13-259(53.88˚S, 4.93˚W; 수심 1,754 m)는 마지막 빙하기부터 퇴빙기까지 생물기원 오팔 함량의 평균값이 39.6%이며, 홀로세 동안 65.9%로 증가한다(Fig. 5d; Kumar et al. 1993).
남극전선 남쪽 해역은 빙하기 동안 생물기원 오팔 생산이 감소했고, 홀로세 동안 온난화되면서 오팔 생산이 증가하는 특징을 보인다. 선행 연구들은 이러한 특징이 나타나는 이유를 해빙 분포 범위의 확장과 증가된 수층의 성층화로 발표하였다(e.g. Charles et al. 1991; Chase et al. 2003; Civel-Mazens et al. 2021; Kaiser et al. 2021). Charles et al. (1991)은 생물기원 오팔 함량을 조절하는 주요한 요인을 해빙면적의 변화로 설명하였고, 남극해에서 빙하기 동안 북쪽으로 확장된 해빙의 영향으로 규조류의 일차생산이 감소했다고 주장하였다. 해빙의 확장 이외에도 빙하기 동안 에크만 수송이 감소하면서 영양염의 용승을 제한하는 수층의 성층화가 강화되어 표층수로 용승을 통한 규산염의 공급이 제한되면서 일차생산이 감소하였다(Francois et al. 1997; Sigman and Boyle 2000; Toggweiler et al. 2006; Kaiser et al. 2021). 한편, De La Rocha et al. (1998)과 Brzezinski et al. (2002)는 수층의 성층화와 더불어 규조류가 일차생산에 용승한 규소를 비효율적으로 활용했을 가능성도 제시하였다.
빙하기 동안 감소한 생물기원 오팔 함량이 퇴빙기 이후 증가하는 양상을 보이는 남극 전선 남쪽 해역과는 반대로, 남극 전선의 북쪽 해역은 빙하기 동안 증가한 생물기원 오팔 함량이 퇴빙기 동안 감소하는 양상을 보인다. 남대서양에 위치한 남극 전선 남쪽의 남미–남극 해령(South America–Antarctic Ridge)에서 획득된 코어 RC15-93(46.10˚S, 13.23˚W; 수심 2,714 m)과 RC13-254(48.57˚S, 5.13˚E; 수심 3,636 m)는 상기한 남극 전선 북쪽 해역의 전형적인 생물기원 오팔 함량 변화를 보인다(Fig. 6; Charles et al. 1991). 코어 RC15-93은 마지막 빙하기 동안 생물기원 오팔 함량의 평균값이 49.9%인 반면, 퇴빙기 동안에는 18.4%로 약 2배 이상 감소한다. 또한, 코어 RC13-254의 생물기원 오팔 함량의 평균값은 56.7%에서 퇴빙기 동안 32.3%로 급격한 감소를 보인다. Charles et al. (1991)은 남극 전선 북쪽에서 빙하기 동안 생물기원 오팔 함량이 증가한 이유를 표층 해수로의 규소 공급이 증가하면서 영양염의 이용 가능성이 커졌기 때문이라고 설명했다. Sigman et al. (1999)은 해빙이 없는 지역에 남극 전선을 가로질러 용해된 규소와 철이 공급되면서 규조류의 일차생산이 증가하였다고 밝혀 이를 뒷받침한다.
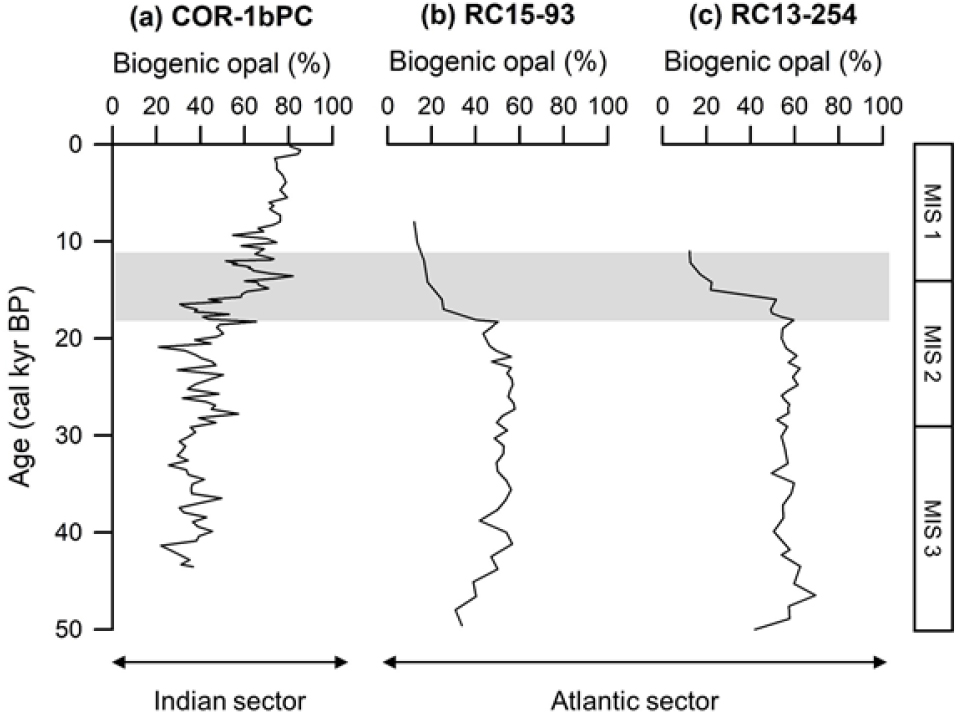
Fig. 6.
Comparison of downcore biogenic opal content between (a) COR-1bPC collected from the south of the Antarctic Polar Front (APF) and (b) RC15-93 and (c) RC13-254 (Charles et al. 1991) collected from the north of the APF
남극 전선 남쪽 해역의 해빙 변화
제4기 후기 남극해의 생물기원 퇴적물의 분포는 남극순환류의 지역적인 특성을 반영하며, 남극 대륙의 빙하 역학, 해빙 분포, 온도 구배, 바람의 특성 및 열염분 순환에 의해 제어된다(Diekmann 2007). 그 중에서도 빛 가용성을 조절하는 해빙의 확장은 남극 전선 남쪽 해역에서 생물기원 오팔 함량이 감소되는 주요 원인들 중 하나이다(Burckle and Cirilli 1987; Chase et al. 2015). 고위도 극지방에만 분포하는 공간적인 한계에도 불구하고 해빙은 지구 기후와 해양 역학에서 중요한 역할을 한다(Zwally et al. 2002; Esper and Gersonde 2014). 해빙은 입사되는 태양 복사의 감소와 같은 직접적인 효과를 통해 생물 펌프에 영향을 미친다. 또한, 간접적으로는 대기–해양 기체 교환의 교란, 그리고 해양에 의존하는 동역학 및 열역학적 프로세스에 영향을 미친다(Anderson et al. 2002). 해빙은 대기–해양의 열교환 상호 작용의 결과로, 해빙의 변화를 이해하는 것은 남극해의 기후 시스템 전반을 이해하는 것과 관련이 있다(Hobbs et al. 2016). 남극해에 해빙이 형성되면 알베도의 계절적인 변화가 발생하여 태양광의 흡수가 달라진다(Ebert et al. 1995). 남극해 표층 수괴의 일차생산성은 빛의 가용도에 의해 조절되기 때문에 일차생산과 내보내기 생산은 해빙의 영향을 받는다. 따라서 남극해의 해빙은 표층 해수로 투과하는 태양광을 차단하여 식물플랑크톤의 성장을 방해하기 때문에 일반적으로 해빙이 확장된 시기에는 표층 해수의 일차생산성이 감소하여 심해로 침강하는 입자 플럭스가 적게 관측된다(e.g. Abelmann and Gersonde 1991; Wefer and Fischer 1991). 코어 COR-1bPC의 오팔 함량 결과와 같이 남극 전선의 남쪽 해역에서 일반적으로 생물기원 오팔 함량은 빙하기에 낮고 간빙기에 높다(Mortlock et al. 1991; Bareille et al. 1998; Chase et al. 2003; Dezileau et al. 2003; Bradtmiller et al. 2009; Sprenk et al. 2013). 이것은 표층 해수의 규조류 생산성이 빙하기 동안 감소하고, 간빙기 동안 증가한다는 것을 의미한다. 이러한 규조류 생산성 변화는 빙하기 동안 해빙의 분포가 확장되어 표층 해수로 투과되는 태양광이 감소하였고 유광층으로 유입되는 햇빛이 제한되어서 표층 해수의 규조류 생산성이 감소한 것으로 해석된다.
남극 대륙 주변부에 가까운 남극 해역에서 빙하기–간빙기 오팔 생산성의 변화는 해빙의 발달과 관련된다(e.g., Charles et al. 1991; Hillenbrand and Cortese 2006; Chase et al. 2015; Tang et al. 2016; Wu and Hou 2017). 남극 대륙에서 획득한 EDC 빙하 코어에서 분석된 Na 플럭스는 지난 74만 년 동안 빙하기 겨울철 해빙의 범위가 두 배로 확장되었음을 지시한다(Wolff et al. 2006). 현재 연구지역인 남극해의 인도양 해역에서 여름철 해빙은 -2°C인 등온선을 따라 분포하며, 이에 따라 65°S에서 70°S 사이에 분포한다(Ferrari et al. 2014). 이에 비해 현재 겨울철 해빙의 가장자리는 ~60°S까지 확장되어 분포한다(Wilson et al. 2019). Benz et al. (2016)은 Fragilariopsis curta와 Fragilariopsis cylindrus의 분포를 바탕으로 마지막 최대 빙하기 동안 남서 인도양 해역에서 현재보다 겨울철 해빙 범위가 60–70% 정도 확장되었다고 밝혔다. 이외에도 다양한 연구에서 규조종 분석에 의하여 빙하기 동안 남극해의 인도양 해역에서 해빙의 최대 범위를 복원하였으며, 해빙의 분포는 빙하기 동안 북쪽으로 더 확장되었다(e.g., Crosta et al. 2004; Gersonde et al. 2005; Chadwick et al. 2020; Ghadi et al. 2020). 이를 통해 빙하기 동안 남극 해역에서 겨울철 해빙의 경계가 50°S까지 확장되어 연구지역의 북쪽까지 표층 해수와 일차생산성에 영향을 미쳤다는 것을 알 수 있다. Orme et al. (2020)은 콘래드 해령에서 획득한 코어 COR-1GC를 이용하여 14 ka 동안의 규조류 군집 특성을 조사하였다. 이 연구결과에 의하면 남극순환류가 우세한 남극해 겨울철 해빙 가장자리와 아남극 전선 사이에서 최대 ~80%까지 나타나는 고유종인 Fragilariopsis kerguelensis가 평균 72.0%로 높게 산출된다(Gersonde and Wefer 1987; Shukla et al. 2013; Orme et al. 2020). Fragilariopsis kerguelensis는 퇴빙기 동안 높은 풍부도를 보이다가 약 10 ka부터 감소한다. 또한, 해빙의 영향이 증가할 때 감소하는 Thalassiosira gracilis는 퇴빙기 동안 평균 1.7%의 풍부도를 보이다가 약 10 ka 이후 평균 2.5%로 산출이 증가한다. 이러한 규조류 군집 분석 결과에 의해 빙하기 동안 콘래드 해령 지역에서도 빙하기 동안 해빙이 증가하여 오팔 생산성이 감소한 것으로 해석된다.
남극전선 남쪽 해역의 수층 안정화 변화
남극해 표층 해수의 일차생산성을 제어하는 또 다른 요인은 온도가 낮고 영양염이 풍부한 해수가 용승하여 표층에 공급되는 미량 영양염의 농도이다(Dezileau et al. 2003). 규조류의 생리학적 상태와 성장 속도는 철에 의해 제한될 수 있지만, 궁극적으로 퇴적되는 생물기원 오팔 함량의 총량은 해수에 용해된 영양염의 공급에 의해 제한된다(Boyd et al. 2000). 현재 남극 전선 남쪽의 높은 생산성은 남극 전선과 겨울철 해빙 가장자리 사이에서 높은 농도의 질산염과 규산염을 표층으로 운반하는 심해의 용승에 의존한다(DeMaster 1981; Manoj and Thamban 2015). 따라서 남극 해역에서 생물기원 오팔의 생산 및 퇴적은 용승 및 수층의 성층화와 직접적인 연관을 갖는다(Anderson et al. 2009). 남극 해역에서 용승은 편서풍의 세기가 약해지거나, 편서풍이 더 북쪽으로 이동했을 때 그 세기가 감소하며, 수직 혼합은 해수의 밀도에 의한 영향을 많이 받는다(Toggweiler et al. 2006; Jaccard et al. 2013). 이외에도 Stephens and Keeling (2000)은 수층의 성층화는 해빙 범위의 증가에 기인했을 가능성이 있다고 주장했다. 극지방 해양의 수직적인 안정성은 일반적으로 온도와 염분의 상호작용에 의해 조절된다(Wilson et al. 2019). Sigman et al. (2004)은 밀도는 낮은 온도에서 온도에 덜 의존적인 경향을 보이며, 이러한 특성으로 인해 해양의 평균 온도가 낮아질수록 표층 해수와 심층 해수 사이의 온도 차이는 더 작은 밀도 차이를 유발하여 염분이 수층 안정화에 더 큰 영향을 미친다고 주장했다. 또한, 해빙이 형성되는 해역에서 겨울철 해빙 범위의 확장은 강력한 수온약층을 유발하는 겨울수괴(Winter Water)를 형성하여 여름철 수층의 성층화를 강화시켰다(Honjo 2004; Hillenbrand and Cortese 2006). 따라서 남극해에서 빙하기 동안 증가된 해빙 면적으로 인해 표층 해수가 냉각되면서 수층의 안정화가 유지되었다(Sigman et al. 2004; Lynch-Stieglitz et al. 2016; Wilson et al. 2019). Francois et al. (1997)은 수층의 안정화로 인해 용승이 약화되어 표층 해수로 공급되는 영양염이 감소하였고, 이로 인해 규조류의 일차생산이 감소했다고 주장했다. 수온과 염분에 의한 성층화 이외에도 Toggweiler et al. (2006)은 마지막 빙하기 동안 남극 상공에서 약해진 서풍이 에크만 수송에 의한 용승을 감소시켜 표층 해양의 영양염 공급이 감소했다고 주장했다. 이와는 반대로 남극 해역에서 바람의 세기가 강해지면 남극순환류에 의한 용승이 증가된다(Marshall and Speer 2012). 이러한 용승은 남극해의 표층 해수에 더 많은 양의 규소를 제공하고, 규조류의 생산에서 제한 요소였던 규소의 공급은 규조류의 생산성을 향상시켜 생물기원 오팔 퇴적의 증가로 이어진다(Sigmon et al. 2002; Bradtmiller et al. 2009).
이와 같은 빙하기–간빙기 오팔 생산성의 변화는 적어도 지난 ~150 kyr까지 반복된다(e.g., Studer et al. 2015; Ghadi et al. 2020). Francois et al. (1997)은 미화석 규조류 군집과 δ13C 값을 이용하여 마지막 빙하기 동안 남극 해역의 남쪽에서 수층이 매우 안정적이었으며, 이와 반대로 남극 해역의 북쪽에서는 수직 혼합이 증가했을 것이라고 주장했다. 마찬가지로 같은 코어에서 Frank et al. (2000)는 10Be을 이용하여 빙하기 동안 수층의 성층화가 표층으로의 영양염 공급을 제한하였을 것이라고 밝혔다. 따라서 남극 해역에서 빙하기 동안 증가된 수층의 성층화는 규조 생산성의 감소로 이어진다는 것을 알 수 있다(Martínez-Garcia et al. 2011). 이러한 연구결과들은 수층의 안정화가 빙하기 동안 규조류의 일차생산성을 조절하여 생산성 감소라는 결과로 이어졌음을 의미한다.
5. 결 론
전 지구적으로 생지화학적 순환에 있어서 중요한 해역인 남극해에서 일차생산자인 규조류에 의한 탄소 순환의 변화를 알아보고자 연구를 수행하였다. 이를 위해 남극해 인도양 해역에서 획득된 코어 COR-1bPC을 이용하여 마지막 빙하기부터 간빙기인 홀로세까지 생물기원 오팔 함량 변화를 통해 남극해 표층 해수의 특성 변화를 복원하였다. 퇴적물의 L*와 대자율은 코어 하부에서 상부로 갈수록 감소한다. 이러한 변화는 코어 하부와 상부 퇴적물을 구성하는 입자와 그 특성이 변화한 것으로 해석된다. 생물기원 오팔 함량은 마지막 빙하기에서 홀로세로 가면서 점차 증가한다. 특히 MIS 2 후반에 30.6%에서 82.0%로 함량이 약 두 배 이상으로 급격하게 증가하며, 이는 남극 빙하에 기록된 기후 인자의 변화(EDC ΔT, AICC CO2 농도)와 일치한다. 이와 같은 생물기원 오팔 함량의 변화가 나타나는 이유는 해빙 범위 확장에 따른 빛 가용성, 해빙의 확장에 따른 수층 안정화로 인한 용승 세기의 변화 때문으로 해석된다. 기존 연구의 규조류 군집 분석 결과를 통해 마지막 빙하기 동안 해빙 범위가 60°S까지 확장되어 콘래드 해령 해역의 규조류 일차생산이 감소된 것으로 해석된다. 또한, 마지막 빙하기 동안 해빙의 확장으로 인한 수온약층의 강화는 수층의 안정화를 유발하여 용승이 약화되었고, 표층으로의 영양염 공급이 제한되어 규조 생산성이 감소한 것으로 해석된다. 이 연구는 비교적 짧은 지질학적 시간 규모의 변화를 복원하였지만, 더 긴 시간 동안의 생물기원 오팔 함량 변화를 이해하기 위한 연구가 필요하다. 그럼에도 불구하고 이 연구결과는 과거 기후 변화 복원에서 남극해 표층 해수의 역할을 이해하기 위한 필수적인 정보를 제공한다.
Acknowledgements
논문을 심사하시고 훌륭한 조언을 해 주신 두 분의 심사위원들게 감사를 드립니다. 이 연구는 부산대학교 기본연구지원사업(2년)에 의하여 수행되었습니다.
References
Abelmann A, Gersonde R (1991) Biosiliceous particle flux in the Southern Ocean. Mar Chem 35(1-4):503-536
10.1016/S0304-4203(09)90040-8Anderson RF, Ali S, Bradtmiller LI, Nielsen SHH, Fleisher MQ, Anderson BE, Burckle LH (2009) Wind-driven upwelling in the Southern Ocean and the deglacial rise in atmospheric CO
2. Science 323(5920):1443-1448
10.1126/science.116744119286547Anderson RF, Chase Z, Fleisher MQ, Sachs J (2002) The Southern Ocean's biological pump during the last glacial maximum. Deep-Sea Res Pt II 49(9-10):1909-1938
10.1016/S0967-0645(02)00018-8Bard E (1988) Correction of accelerator mass spectrometry
14C ages measured in planktonic foraminifera: paleoceanographic implications. Paleoceanography 3(6):635-645
10.1029/PA003i006p00635Bareille G, Labracherie M, Bertrand P, Labeyrie L, Lavaux G, Dignan M (1998) Glacial-interglacial changes in the accumulation rates of major biogenic components in Southern Indian Ocean sediments. J Marine Syst 17(1-4):527-539
10.1016/S0924-7963(98)00062-1Batchelor CL, Margold M, Krapp M, Murton DK, Dalton AS, Gibbard PL, Stokes CR, Murton JB, Manica A (2019) The configuration of Northern Hemisphere ice sheets through the Quaternary. Nat Comm 10(1):3713
10.1038/s41467-019-11601-2Belkin IM, Gordon AL (1996) Southern Ocean fronts from the Greenwich meridian to Tasmania. J Geophys Res-Oceans 101(C2):3675-3696
10.1029/95JC02750Benz V, Esper O, Gersonde R, Lamy F, Tiedemann R (2016) Last Glacial Maximum sea surface temperature and sea-ice extent in the Pacific sector of the Southern Ocean. Quaternary Sci Rev 146:216-237
10.1016/j.quascirev.2016.06.006Bereiter B, Eggleston S, Schmitt J, Nehrbass‐Ahles C, Stocker TF, Fischer H, Kipfstuhl S, Chappellaz J (2015) Revision of the EPICA Dome C CO
2 record from 800 to 600 kyr before present. Geophys Res Lett 42(2):542-549
10.1002/2014GL061957Berger A (1988) Milankovitch theory and climate. Rev Geophys 26(4):624-657
10.1029/RG026i004p00624Boyd PW, Watson AJ, Law CS, Abraham ER, Trull T, Murdoch R, Bakker DCE, Bowie AR, Buesseler KO, Chang H, Charette M, Croot P, Downing K, Frew R, Gall M, Hadfield M, Hall J, Harvey M, Jameson G, LaRoche J, Liddicoat M, Maldonado MT, McKay RM, Nodder S, Pickmere S, Pridmore R, Rintoul S, Safi K, Sutton P, Strzepek R, Tanneberger K, Turner S, Waite A, Zeldis J (2000) A mesoscale phytoplankton bloom in the polar Southern Ocean stimulated by iron fertilization. Nature 407(6805):695-702
10.1038/3503750011048709Bradtmiller LI, Anderson RF, Fleisher MQ, Burckle LH (2009) Comparing glacial and Holocene opal fluxes in the Pacific sector of the Southern Ocean. Paleoceanography 24(2):PA2214
10.1029/2008PA001693Brzezinski MA, Pride CJ, Franck VM, Sigman DM, Sarmiento JL, Matsumoto K, Gruber N, Rau GH, Coale KH (2002) A switch from Si(OH)
4 to NO
3− depletion in the glacial Southern Ocean. Geophys Rese Lett 29(12):5-1-5-4
10.1029/2001GL014349Burckle LH, Cirilli J (1987) Origin of diatom ooze belt in the Southern Ocean; implications for late Quatenary paleoceanography. Micropaleontology 33(1):82-86
10.2307/1485529Chadwick M, Allen CS, Sime LC, Hillenbrand CD (2020) Analysing the timing of peak warming and minimum winter sea-ice extent in the Southern Ocean during MIS 5e. Quaternary Sci Rev 229:106134
10.1016/j.quascirev.2019.106134Charles CD, Froelich PN, Zibello MA, Mortlock RA, Morley JJ (1991) Biogenic opal in Southern Ocean sediments over the last 450,000 years: implications for surface water chemistry and circulation. Paleoceanography 6(6):697-728
10.1029/91PA02477Chase Z, Anderson RF, Fleisher MQ, Kubik PW (2003) Accumulation of biogenic and lithogenic material in the Pacific sector of the Southern Ocean during the past 40,000 years. Deep-Sea Res Pt II 50(3-4):799-832
10.1016/S0967-0645(02)00595-7Chase Z, Kohfeld KE, Matsumoto K (2015) Controls on biogenic silica burial in the Southern Ocean. Global Biogeochem Cy 29(10):1599-1616
10.1002/2015GB005186Civel-Mazens M, Crosta X, Cortese G, Michel E, Mazaud A, Ther O, Ikehara M, Itaki T (2021) Antarctic Polar Front migrations in the Kerguelen Plateau region, Southern Ocean, over the past 360 kyrs. Global Planet Change 202:103526
10.1016/j.gloplacha.2021.103526Cooke DW, Hays JD (1982) Estimates of Antarctic Ocean seasonal sea-ice cover during glacial intervals. Antarct Geosci 131:1017-1025
Cortese G, Gersonde R, Hillenbrand CD, Kuhn G (2004) Opal sedimentation shifts in the World Ocean over the last 15 Myr. Earth Planet Sci Lett 224(3-4):509-527
10.1016/j.epsl.2004.05.035Crosta X, Romero O, Armand LK, Pichon JJ (2005) The biogeography of major diatom taxa in Southern Ocean sediments: 2. Open ocean related species. Palaeogeogr Palaeocl 223(1-2):66-92
10.1016/j.palaeo.2005.03.028Crosta X, Shemesh A, Salvignac ME, Gildor H, Yam R (2002) Late Quaternary variations of elemental ratios (C/Si and N/Si) in diatom-bound organic matter from the Southern Ocean. Deep-Sea Res Pt II 49(9-10):1939-1952
10.1016/S0967-0645(02)00019-XCrosta X, Sturm A, Armand L, Pichon JJ (2004) Late Quaternary sea ice history in the Indian sector of the Southern Ocean as recorded by diatom assemblages. Mar Micropaleontol 50(3-4):209-223
10.1016/S0377-8398(03)00072-0De La Rocha CL, Brzezinski MA, DeNiro MJ, Shemesh A (1998) Silicon-isotope composition of diatoms as an indicator of past oceanic change. Nature 395(6703):680-683
10.1038/27174DeMaster DJ (1981) The supply and accumulation of silica in the marine environment. Geochim Cosmochim Ac 45(10):1715-1732
10.1016/0016-7037(81)90006-5Dezileau L, Reyss JL, Lemoine F (2003) Late Quaternary changes in biogenic opal fluxes in the Southern Indian Ocean. Mar Geol 202(3-4):143-158
10.1016/S0025-3227(03)00283-4Diekmann B (2007) Sedimentary patterns in the late Quaternary Southern Ocean. Deep-Sea Res Pt II 54(21-22):2350-2366
10.1016/j.dsr2.2007.07.025Durgadoo JV, Lutjeharms JRE, Biastoch A, Ansorge IJ (2008) The Conrad Rise as an obstruction to the Antarctic Circumpolar Current. Geophy Res Lett 35(20):L20606
10.1029/2008GL035382Ebert EE, Schramm JL, Curry JA (1995) Disposition of solar radiation in sea ice and the upper ocean. J Geophys Res-Oceans 100(C8):15965-15975
10.1029/95JC01672Esper O, Gersonde R (2014) New tools for the reconstruction of Pleistocene Antarctic sea ice. Palaeogeogr Palaeocl 399:260-283
10.1016/j.palaeo.2014.01.019Ferrari R, Jansen MF, Adkins JF, Burke A, Stewart AL Thompson AF (2014) Antarctic sea ice control on ocean circulation in present and glacial climates. P Natl Acad Sci USA 111(24):8753-8758
10.1073/pnas.13239221114066517Francois R, Altabet MA, Yu EF, Sigman DM, Bacon MP, Frank M, Bohrmann G, Bareille G, Labeyrie LD (1997) Contribution of Southern Ocean surface-water stratification to low atmospheric CO
2 concentrations during the last glacial period. Nature 389(6654):929-935
10.1038/40073Frank M, Gersonde R, Loeff MR, Bohrmann G, Nürnberg CC, Kubik PW, Suter M, Mangini A (2000) Similar glacial and interglacial export bioproductivity in the Atlantic sector of the Southern Ocean: multiproxy evidence and implications for glacial atmospheric CO
2. Paleoceanography 15(6):642-658
10.1029/2000PA000497Gersonde R, Crosta X, Abelmann A, Armand L (2005) Sea-surface temperature and sea ice distribution of the Southern Ocean at the EPILOG Last Glacial Maximum-a circum - Antarctic view based on siliceous microfossil records. Quaternary Sci Rev 24(7-9):869-896
10.1016/j.quascirev.2004.07.015Gersonde R, Wefer G (1987) Sedimentation of biogenic siliceous particles in Antarctic waters from the Atlantic sector. Mar Micropaleontol 11(4):311-332
10.1016/0377-8398(87)90004-1Ghadi P, Nair A, Crosta X, Mohan R, Manoj MC, Meloth T (2020) Antarctic sea-ice and palaeoproductivity variation over the last 156,000 years in the Indian sector of Southern Ocean. Mar Micropaleontol 160:101894
10.1016/j.marmicro.2020.101894Gordon AL (1971) Antarctic polar front zone. Antarctic Oceanol 15:205-221
10.1029/AR015p0205Gruber N, Gloor M, Mikaloff Fletcher SE, Doney SC, Dutkiewicz S, Follows MJ, Gerber M, Jacobson AR, Joos F, Lindsay K, Menemenlis D, Mouchet A, Müller SA, Sarmiento JL, Takahashi T (2009) Oceanic sources, sinks, and transport of atmospheric CO
2. Global Biogeochem Cy 23(1):GB1005
10.1029/2008GB003349Henley SF, Cavan EL, Fawcett SE, Kerr R, Monteiro T, Sherrell RM, Bowie AR, Boyd PW, Barnes DKA, Schloss IR, Marshall T, Flynn R, Smith S (2020) Changing biogeochemistry of the Southern Ocean and its ecosystem implications. Front Mar Sci 7:581
10.3389/fmars.2020.00581Hillenbrand CD, Cortese G (2006) Polar stratification: a critical view from the Southern Ocean. Palaeogeogr Palaeocl 242(3-4):240-252
10.1016/j.palaeo.2006.06.001Hobbs WR, Massom R, Stammerjohn S, Reid P, Williams G, Meier W (2016) A review of recent changes in Southern Ocean sea ice, their drivers and forcings. Global Planet Change 143:228-250
10.1016/j.gloplacha.2016.06.008Honjo S (2004) Particle export and the biological pump in the Southern Ocean. Antarc Sci 16(4):501-516
10.1017/S0954102004002287Jaccard SL, Hayes CT, Martinez-Garcia A, Hodell DA, Anderson RF, Sigman DM, Haug GH (2013) Two modes of change in Southern Ocean productivity over the past million years. Science 339(6126):1419-1423
10.1126/science.122754523520109Jouzel J, Masson-Delmotte V, Cattani O, Dreyfus G, Falourd S, Hoffmann G, Minster B, Nouet J, Barnola JM, Chappellaz J, Fischer H, Gallet JC, Johnsen S, Leuenberger M, Loulergue L, Luethi D, Oerter H, Parrenin F, Raisbeck G, Raynaud D, Schilt A, Schwander J, Selmo E, Souchez R, Spahni R, Stauffer B, Steffensen JP, Stenni B, Stocker TF, Tison JL, Werner M, Wolff EW (2007) Orbital and millennial Antarctic climate variability over the past 800,000 years. Science 317(5839):793-796
10.1126/science.114103817615306Kaiser EA, Billups K, Bradtmiller L (2021) A 1 million year record of biogenic silica in the Indian Ocean sector of the Southern Ocean: regional versus global forcing of primary productivity. Paleoceanography and Paleoclimatology 36(3):e2020PA004033
10.1029/2020PA004033Kohfeld KE, Quéré CL, Harrison SP, Anderson RF (2005) Role of marine biology in glacial-interglacial CO
2 cycles. Science 308(5718):74-78
10.1126/science.110537515802597Köhler P, Fischer H, Munhoven G, Zeebe RE (2005) Quantitative interpretation of atmospheric carbon records over the last glacial termination. Global Biogeochem Cy 19(4):GB4020
10.1029/2004GB002345Kumar N, Anderson RF, Mortlock RA, Froelich PN, Kubik P, Dittrich-Hannen B, Suter M (1995) Increased biological productivity and export production in the glacial Southern Ocean. Nature 378(6558):675-680
10.1038/378675a0Kumar N, Gwiazda R, Anderson RF, Froelich PN (1993)
231Pa/
230Th ratios in sediments as a proxy for past changes in Southern Ocean productivity. Nature 362(6415):45-48
10.1038/362045a0Latimer JC (2004) Paleo-export production and terrigenous sedimentation in the Southern Ocean. Ph. D. Thesis, Indiana University, 29 p
Lazarus D, Caulet JP (1993) Cenozoic Southern Ocean reconstructions from sedimentologic, radiolarian, and other microfossil data. In: Kennett P, Warnke DA (eds) The Antarctic Paleoenvironment: a perspective on global change. American Geophysical Union, pp 145-174
Lynch-Stieglitz J, Ito T, Michel E (2016) Antarctic density stratification and the strength of the circumpolar current during the Last Glacial Maximum. Paleoceanography 31(5):539-552
10.1002/2015PA002915Manoj MC, Thamban M (2015) Shifting frontal regimes and its influence on bioproductivity variations during the Late Quaternary in the Indian sector of Southern Ocean. Deep-Sea Res Pt II 118:261-274
10.1016/j.dsr2.2015.03.011Marinov I, Gnanadesikan A, Toggweiler JR, Sarmiento JL (2006) The southern ocean biogeochemical divide. Nature 441(7096):964-967
10.1038/nature0488316791191Marshall J, Speer K (2012) Closure of the meridional overturning circulation through Southern Ocean upwelling. Nat Geosci 5(3):171-180
10.1038/ngeo1391Martin JH (1990) Glacial‐interglacial CO
2 change: the iron hypothesis. Paleoceanography 5(1):1-13
10.1029/PA005i001p00001Martínez-Garcia A, Rosell‐Melé A, Geibert W, Gersonde R, Masqué P, Gaspari, V, Barbante C (2009) Links between iron supply, marine productivity, sea surface temperature, and CO
2 over the last 1.1 Ma. Paleoceanography 24(1):PA1207
10.1029/2008PA001657Martínez-Garcia A, Rosell-Melé A, Jaccard SL, Geibert W, Sigman DM, Haug GH (2011) Southern Ocean dust-climate coupling over the past four million years. Nature 476(7360):312-315
10.1038/nature1031021814203Moore JK, Abbott MR, Richman JG, Nelson DM (2000) The Southern Ocean at the last glacial maximum: a strong sink for atmospheric carbon dioxide. Global Biogeochem Cy 14(1):455-475
10.1029/1999GB900051Mortlock RA, Charles CD, Froelich PN, Zibello MA, Saltzman J, Hays JD, Burckle LH (1991) Evidence for lower productivity in the Antarctic Ocean during the last glaciation. Nature 351(6323):220-223
10.1038/351220a0Mortlock RA, Froelich PN (1989) A simple method for the rapid determination of biogenic opal in pelagic marine sediments. Deep-Sea Res 36(9):1415-1426
10.1016/0198-0149(89)90092-7Nielsen SH, Hodell DA, Kamenov G, Guilderson T, Perfit MR (2007) Origin and significance of ice‐rafted detritus in the Atlantic sector of the Southern Ocean. Geochem Geophy Geosy 8(12):C001618
10.1029/2007GC001618Oiwane H, Ikehara M, Suganuma Y, Miura H, Nakamura Y, Sato T, Nogi Y, Yamane M, Yokoyama Y (2014) Sediment waves on the Conrad Rise, Southern Indian Ocean: implications for the migration history of the Antarctic Circumpolar Current. Mar Geol 348:27-36
10.1016/j.margeo.2013.10.008Orme LC, Crosta X, Miettinen A, Divine DV, Husum K, Isaksson E, Wacker L, Mohan R, Ther O, Ikehara M (2020) Sea surface temperature in the Indian sector of the Southern Ocean over the Late Glacial and Holocene. Clim Past 16(4):1451-1467
10.5194/cp-16-1451-2020Orsi AH, Whitworth T, Nowlin Jr WD (1995) On the meridional extent and fronts of the Antarctic Circumpolar Current. Deep-Sea Res Pt I 42(5):641-673
10.1016/0967-0637(95)00021-WPahnke K, Sachs JP (2006) Sea surface temperatures of southern midlatitudes 0-160 kyr BP. Paleoceanography 21(2):PA2003
10.1029/2005PA001191Paillard D (2001) Glacial cycles: toward a new paradigm. Rev Geophys 39(3):325-346
10.1029/2000RG000091Petit JR, Jouzel J, Raynaud D, Barkov NI, Barnola JM, Basile I, Bender M, Chappellaz J, Davis M, Delaygue G, Delmotte M, Kotlyakov VM, Legrand M, Lipenkov VY, Lorius C, PÉpin L, Ritz C, Saltzman E, Stievenard M (1999) Climate and atmospheric history of the past 420,000 years from the Vostok ice core, Antarctica. Nature 399(6735):429-436
10.1038/20859Petrou K, Kranz SA, Trimborn S, Hassler CS, Ameijeiras SB, Sackett O, Davidson AT (2016) Southern Ocean phytoplankton physiology in a changing climate. J Plant Physiol 203:135-150
10.1016/j.jplph.2016.05.00427236210Pickering KT, Hiscott RN (2015) Deep marine systems: processes, deposits, environments, tectonics and sedimentation. John Wiley & Sons, Hoboken, USA, 485 p
Pollard RT, Lucas MI, Read JF (2002) Physical controls on biogeochemical zonation in the Southern Ocean. Deep-Sea Res Pt II 49(16):3289-3305
10.1016/S0967-0645(02)00084-XRagueneau O, Tréguer P, Leynaert A, Anderson RF, Brzezinski MA, DeMaster DJ, Dugdale RC, Dymond J, Fisher G, Francois R, Heinze C, Maier-Reimer E, Martin-Jezequel V, Nelson DW, Quéguiner B (2000) A review of the Si cycle in the modern ocean: recent progress and missing gaps in the application of biogenic opal as a paleoproductivity proxy. Global Planet Change 26(4):317-365
10.1016/S0921-8181(00)00052-7Rotzien JR, Hernández-Molina FJ, Fonnesu M, Thieblemont A (2022) Deepwater sedimentation units. In Deepwater Sedimentary Systems. Elsevier, Amsterdam, 203 p
10.1016/B978-0-323-91918-0.00009-8Shukla SK, Crosta X, Cortese G, Nayak GN (2013) Climate mediated size variability of diatom
Fragilariopsis kerguelensis in the Southern Ocean. Quaternary Sci Rev 69:49-58
10.1016/j.quascirev.2013.03.005Siddall M, Rohling EJ, Almogi-Labin A, Hemleben C, Meischner D, Schmelzer I, Smeed DA (2003) Sea-level fluctuations during the last glacial cycle. Nature 423(6942):853-858
10.1038/nature0169012815427Sigman DM, Altabet MA, Francois R, McCorkle DC, Gaillard JF (1999) The isotopic composition of diatom-bound nitrogen in Southern Ocean sediments. Paleoceanography, 14(2):118-134
10.1029/1998PA900018Sigman DM, Boyle EA (2000) Glacial/interglacial variations in atmospheric carbon dioxide. Nature 407(6806):859-869
10.1038/3503800011057657Sigman DM, Jaccard SL, Haug GH (2004) Polar ocean stratification in a cold climate. Nature 428(6978):59-63
10.1038/nature0235714999278Sigmon DE, Nelson DM, Brzezinski MA (2002) The Si cycle in the Pacific sector of the Southern Ocean: seasonal diatom production in the surface layer and export to the deep sea. Deep-Sea Res Pt II 49(9-10):1747-1763
10.1016/S0967-0645(02)00010-3Sprenk D, Weber ME, Kuhn G, Rosén P, Frank M, Molina-Kescher M, Liebetrau V, Röhling HG (2013) Southern Ocean bioproductivity during the last glacial cycle-new detection method and decadal-scale insight from the Scotia Sea. In: Mabrey MJ, Barker PF, Barrett PJ, Bowman V, Davies B, Smellie JL, Tranter M (eds) Antarctic palaeoenvironments and earth-surface processes. Geological Society of London, pp 245-261
10.1144/SP381.17Stephens BB, Keeling RF (2000) The influence of Antarctic sea ice on glacial-interglacial CO
2 variations. Nature 404(6774):171-174
10.1038/3500455610724166Studer AS, Sigman DM, Martínez‐García A, Benz V, Winckler G, Kuhn G, Esper O, Lamy F, Jaccard SL, Wacker L, Oleynik S, Gersonde R, Haug GH (2015) Antarctic Zone nutrient conditions during the last two glacial cycles. Paleoceanography 30(7):845-862
10.1002/2014PA002745Stuiver M, Reimer PJ (1993) Extended
14C data base and revised CALIB 3.0
14C age calibration program. Radiocarbon 35(1):215-230
10.1017/S0033822200013904Tang Z, Shi X, Zhang X, Chen Z, Chen MT, Wang X, Wang H, Liu H, Lohmann G, Li P, Ge S, Huang Y (2016) Deglacial biogenic opal peaks revealing enhanced Southern Ocean upwelling during the last 513 ka. Quatern Int 425:445-452
10.1016/j.quaint.2016.09.020Thamban M, Naik SS, Mohan R, Rajakumar A, Basavaiah N, D'Souza W, Sarita Kerkar, Subramaniam MM, Sudhakar M, Pandey PC (2005) Changes in the source and transport mechanism of terrigenous input to the Indian sector of Southern Ocean during the late Quaternary and its palaeoceanographic implications. J Earth Syst Sci 114:443-452
10.1007/BF02702021Toggweiler JR, Russell JL, Carson SR (2006) Midlatitude westerlies, atmospheric CO
2, and climate change during the ice ages. Paleoceanography 21(2):PA2005
10.1029/2005PA001154Treguer P, Nelson DM, Van Bennekom AJ, DeMaster DJ, Leynaert A, Quéguiner B (1995) The silica balance in the world ocean: a reestimate. Science 268(5209):375-379
10.1126/science.268.5209.37517746543Veres D, Bazin L, Landais A, Toyé-Mahamadou-Kele H, Lemieux-Dudon B, Parrenin F, Martinerie P, Blayo E, Blunier T, Capron E, Chappellaz J, Rasmussen SO, Severi M, Svensson A, Vinther B, Wolff EW (2013) The Antarctic ice core chronology (AICC2012): an optimized multi-parameter and multi-site dating approach for the last 120 thousand years. Clim Past 9(4):1733-1748
10.5194/cp-9-1733-2013Wefer G, Fischer G (1991) Annual primary production and export flux in the Southern Ocean from sediment trap data. Mar Chem 35(1-4):597-613
10.1016/S0304-4203(09)90045-7Whitworth T, Nowlin Jr WD (1987) Water masses and currents of the Southern Ocean at the Greenwich Meridian. J Geophys Res-Oceans 92(C6):6462-6476
10.1029/JC092iC06p06462Wilson EA, Riser SC, Campbell EC, Wong AP (2019) Winter upper-ocean stability and ice-ocean feedbacks in the sea ice-covered Southern Ocean. J Phys Oceanogr 49(4):1099-1117
10.1175/JPO-D-18-0184.1Wolff EW, Fischer H, Fundel F, Ruth U, Twarloh B, Littot GC, Mulvaney R, Röthlisberger R, de Angelis M, Boutron CF, Hansson M, Jonsell U, Hutterli MA, Lambert F, Kaufmann P, Stauffer B, Stocker TF, Steffensen JP, Bigler M, Siggaard-Andersen ML, Udisti R, Becagli S, Castellano E, Severi M, Wagenbach D, Barbante C, Gabrielli P, Gaspari V (2006) Southern Ocean sea-ice extent, productivity and iron flux over the past eight glacial cycles. Nature 440(7083):491-496
10.1038/nature0461416554810Wu SY, Hou S (2017) Impact of icebergs on net primary productivity in the Southern Ocean. Cryosphere 11(2):707-722
10.5194/tc-11-707-2017Zwally HJ, Comiso JC, Parkinson CL, Cavalieri DJ, Gloersen P (2002) Variability of Antarctic sea ice 1979-1998. J Geophys Res-Oceans 107(C5):9
10.1029/2000JC000733

