

1. 서 론
2. 재료 및 방법
실험생물 채집 및 관리
유주자 방출 유도
수온별 pH 및 염분 농도별 유주자 발아율 실험
자료분석
3. 결 과
수온과 pH 농도별 실험생물의 발아율 변화
수온과 염분 농도별 실험생물의 발아율 변화
수온, pH 및 염분 농도별 실험생물의 민감도 비교
4. 고 찰
5. 적 요
1. 서 론
전 세계적으로 기후변화로 인해 해수의 수온은 지속적으로 상승하고 있으며, 해양 열파(marine heat waves, MHWs)의 지속시간, 빈도 및 강도는 점차적으로 증가하고 있다(Pastor et al. 2018; Bulgin et al. 2020). 이와 같은 해양의 온난화(ocean warming)는 해조류의 종조성, 분포, 생식, 생존 및 대사과정에 영향을 주는 것으로 보고되었다(Wernberg et al. 2016; Khan et al. 2018; Wang et al. 2021). 예를 들면, 일정 수온 범위 내에서 수온 증가는 아미노산, 지방산 및 지질과 같은 화학적 조성을 변화시키며, 효소 활동이 촉진되고, 대사 활동이 활발하게 진행되어 광합성 및 성장에 도움을 줄 수 있으나 임계 수온(critical thermal)을 초과하면 개체군이 감소하거나 사멸의 위험성이 증가하고, 심한 경우 종의 멸종이 가능한 것으로 보고되었다(Gao et al. 2018; Wu et al. 2019).
해양산성화(ocean acidification, OA)는 인위적인 CO2 배출 증가로 인해 탄산염 순환 체계를 변화시켜 pH를 감소시킴으로써 생태계 변화를 유도하고 있다(Feely et al. 2004). 이러한 현상은 해양생물의 다양성과 생태계 기능에 긍정적 또는 부정적 영향을 줄 것으로 예측되었는데, OA는 해조류의 성장과 광합성을 촉진하는 결과를 보였으나 OA에 대한 해조류의 반응은 생물 종 및 발달 단계에 따라 차이를 보일 수 있다(Choi et al. 2019; Gao et al. 2019a, 2019b). 한편 해빙의 녹음, 강우 및 육상으로부터 해양으로 유입되는 담수는 연안 지역에 저 염분 환경을 조성함으로써 해조류의 대사, 성장 및 생존에 영향을 미치는 것으로 보고되었다(Tresnati et al. 2021; Bews et al. 2021). 예를 들면 저염분에 노출된 다시마(Saccharina japonica)는 성장 속도와 색소 감소가 유발되며, 이러한 현상은 세포 내 이온 교환이 원활하지 못하기 때문에 광합성과 영양염 흡수를 통한 생물의 생식, 성장 및 분포 등의 생리-생태 활성에 부정적인 영향을 미치는 것으로 보고되었다(Raybaud et al. 2013; Kumar et al. 2014; Bollen et al. 2016). 한편 최근까지 진행된 해조류의 생태-생리 연구는 수온, 빛, 영양염 등을 중요한 요인으로 선정하여 수행되어 왔으며, 해수에서 강한 완충작용을 하는 pH와 일부 지역에서 일시적으로 변동하는 염분 변화에 노출된 생물의 영향 평가는 상대적으로 크게 주목받지 못하였다(Gaitán-Espitia et al. 2014). 또한 대부분의 연구는 단일 항목에 대한 생물의 성장, 광합성 및 생리 대사 활성을 비교한 연구를 주로 수행되었으나, 해조류의 초기 생활사 단계에서 2종 이상의 복합적인 환경요인의 상호작용에 대한 민감도를 비교 분석한 연구는 미흡한 상황이다.
해양에 서식하는 갈조류 중 미역(Undaria pinnatifida)은 일년생 종으로 조하대 5.0 m 이내에 주로 서식하며, 한국과 일본에서는 외부형태와 상대 생장률의 차이에 따라 북방형과 남방형의 2개 품종으로 구분된다(Okamura 1915). 또한 미역은 봄철에 포자를 방출하는데, 5.0–25.0 °C에서 정상 발아하여 배우체를 거쳐 아포체 단계로 발달한다(Kim and Nam 1997). 넓미역(Undariopsis peterseniana)은 일년생이며, 6월부터 7월까지 성숙한 유주자가 방출되는 종이다(Hwang et al. 2010b). 본 종은 제주도를 포함한 남해안 일부 지역 및 울릉도 조하대 20.0 m 수심보다 깊은 곳에서 광범위한 군락지가 발견되었다(Yoon 2015). 한편 대황(Ecklonia bicyclis)은 다년생 종으로써 경상북도 영덕, 울릉도와 독도 연안 수심 10.0 m 전후의 조하대 지역에 주로 최적 성숙 수온은 15.0–20.0°C로 보고되었다(Kang et al. 2001; Lee and Kim 2020). 미역, 넓미역 및 대황은 대형 갈조류로 분류되며, 이형세대교번을 하고, 배우체 세대에서 암수 배우체가 분리되는 생활사를 가지고 있어 실험실 내에서도 배우체의 배양이 가능한 종이다(Lee and Kim 2020).
생물검정은 다양한 물질 또는 환경인자에 대한 생물의 영향을 직․간접적으로 평가하는 방법이며, 미국과 EU 국가를 포함한 많은 국가에서는 공정시험법으로 제정하여 유해물질 및 환경인자의 위해성을 평가하고 있다(APHA 1995; ISO 1995; ASTM 1996; NIWA 1998; USEPA 2002). 최근 국내에서는 건강한 해양생태계 구현에 따른 수산자원회복이라는 국가적 패러다임의 변화에 부합하여 환경평가 및 물질의 위해성 평가방법으로 다양한 분류군을 해양생물공정시험기준으로 등록하여 연안 환경의 평가도구로 활용하고 있다(MOF 2018). 한편 해양생물공정시험기준에 등재된 해조류는 대형 갈조류인 미역(Undaria pinnatifida)과 다시마(Saccharina japonica)이며, 생물검정은 유주자 발아율, 배우체와 아포체 생장율을 endpoint로 설정하여 해양생태독성평가를 수행하고 있다(MOF 2018). 실험생물은 연중 평가가 가능하도록 암․수 배우체를 분리하고 계대 배양하여 암수배우체 생존 및 상대 성장률 실험에 사용하였는데, 배우체를 이용한 평가방법은 실험실에서 배양된 생물을 대상으로 하기 때문에 전문적인 관리가 필요하다(Lee et al. 2022). 이와 반면 유주자 발아 실험은 현장 채집 후 수 시간 이내에 실시할 수 있으며, 현미경 상에서 관찰이 용이하여 실험생물의 단기 스트레스 영향을 평가할 수 있는 장점이 있다.
국내에서 대형 갈조류를 대상으로 수행된 생물검정 연구는 미역 포자와 암배우체를 이용한 방오도료 성분의 독성평가(Lee 2021), 생태독성평가를 위한 미역 암배우체 생존율 및 상대성장률의 환경조건 연구(Lee et al. 2022), 넓미역과 대황의 초기 생활사에 미치는 pH 및 염분 영향에 관한 연구(Yoon 2023) 및 대황 배우체와 아포체의 생장에 미치는 환경 인자의 영향에 관한 연구(Lee and Kim 2020)가 이루어진 바 있다. 이와 같은 연구는 대부분 수산자원 활성화 및 바다숲 조성용 종자의 대량생산을 위한 기초 연구로써 수온, 광량 및 광합성 효율 등에 관한 기초자료를 확보하기 위해 수행되었다. 또한 해조류의 생태-생리 연구를 통해 기후변화, 해양산성화 및 갯녹음에 따른 연안 해조숲의 생태계 변동 원인을 규명하기 위한 연구는 진행되고 있으나 생활사(life cycle)가 유사하지만 서식환경과 생존 기간이 다른 상기의 대형 갈조류 3종을 대상으로 이들의 초기 생활사에 미치는 2종 이상의 복합적인 환경인자에 노출된 생물의 영향을 규명한 연구는 미흡한 상태이다. 이에 따라 본 연구에는 해양환경 변화에 따른 해조류의 생태-생리 변화를 예측하고자 서식환경 및 생존 기간이 다른 미역, 넓미역 및 대황 등 3종의 대형 갈조류를 대상으로 해양생태독성평가를 통해 다양한 수온과 pH 및 염분이 조합된 환경조건에서 초기 생활사 단계 중 실험생물별 유주자의 발아율 차이를 규명하고, 생물의 생태 반응 및 실험생물별 민감도를 비교 분석하였다.
2. 재료 및 방법
실험생물 채집 및 관리
본 연구에 사용된 미역, 넓미역 및 대황은 경상북도 울릉군 현포리(37°31′N, 130°49′E) 주변 조하대에 서식하는 성숙한 개체를 사용하였는데, 미역의 경우 2024년 5월에 수심 5.0 m, 넓미역은 7월에 25.0 m, 대황은 11월에 5.0 m에서 채집하였다. 각각의 생물은 채집 즉시 30분 이내에 500 m 거리에 위치한 실험실로 옮겨 12.0±1.0°C, 31.0±1.0 psu, pH 8.0±0.2, 산소포화농도 90.0% 이상의 수질환경(YSI, 556MPS, USA)과 어두운 상태를 유지하면서 실험 전까지 유주자 방출을 최소화하였다.
유주자 방출 유도
대형 갈조류 3종의 유주자 방출(미역은 포자엽, 넓미역과 대황의 경우 자낭반)은 0.45 µm GF/C로 여과한 자연 해수를 멸균 처리한 후 15.0±1.0°C 수온에서 유도하였다. 유주자 방출은 성숙한 개체의 포자엽 및 엽체 표면에 부착한 물질을 해수로 제거하고, 거즈(gauze)로 물기를 제거한 후 60분 동안 음건한 상태로 건조 스트레스를 주었다. 건조된 포자엽과 엽체는 100 g을 계량하여 1,000 mL 비이커로 옮기고, 멸균 해수 500 mL를 넣고 약하게 저어 10분 동안 유주자 방출을 유도하였다. 본 연구에 500 mL 멸균 해수에 방출된 유주자는 마이크로 피펫을 사용하여 1 µL를 채취하여 슬라이드 위에 접종한 후 영상처리장치가 장착된 광학현미경(×20)(CKX41, Olympus, Japan) 하에서 100개체/µL 이상 방출된 유주자 개체수를 확인한 후 시험생물로 사용하였다(Fig. 1).

수온별 pH 및 염분 농도별 유주자 발아율 실험
본 연구는 다양한 수온환경에서 각각의 pH 및 염분 농도별 미역, 넓미역 및 대황 유주자의 발아율 실험을 실시하였다. 실험은 항온실 내에서 비교환 정수방식(static non-renewal test)으로 실시하였으며, 실험용수는 0.45 µm GF/C로 여과한 멸균 해수를 사용하였다. 수온은 시료 채집 정점 주변의 연중 수온분포를 고려하여 5.0, 10.0, 15.0, 20.0, 25.0, 30.0°C로 설정하였다. 유주자 배양용기(24-well plate)는 배양액의 수온 차이를 최소화하기 위하여 무작위로 배치하였다. 한편 본 연구는 단기간(72시간) 동안 유주자의 급성 생태 반응을 관찰하기 위해 실험용액의 농도 구간을 광범위하게 세팅하였으며, 멸균 해수(pH 8.0±0.2, 31.0±1.0 psu)을 대조구로 설정하고 순차적으로 희석하여 농도를 조제하였다. 실험용액의 pH는 다항목 수질측정기(HQ400, HACH company, USA)를 사용하여 해수에 염산(HCl)을 넣어 pH 4.0±0.2를 만든 후 pH 8.0 해수를 넣어 단계적으로 희석하여 pH 8.0(대조구), 7.5, 7.0, 6.5, 6.0, 5.5, 5.0, 4.5 농도로 조제하였다. 염분은 염분측정기(YSI 3100, YSI Inc., USA)를 사용하여 멸균 해수에 1차 증류수를 넣어 25.0, 20.0, 15.0, 10.0, 5.0, 0.0 psu까지 희석하여 농도를 조절하였다. 실험은 72시간 동안 수행하였으며, 농도별 4개의 반복구를 주었다. 실험 기간 중 광도는 50.0 µmol photon m-2s-1 상태를 유지하였으며, 광주기는 12:12h L:D (Light:Dark)로 세팅하였다. 실험용액은 24-well plate에 노출 농도별로 3.0 mL를 채웠으며, 각각의 용기에 유주자 현탁액을 10㎕씩 넣고 실험하였다. 실험기간 동안 용액은 교체하지 않았다. 본 실험의 측정요소(endpoint)는 72시간 발아율(spore germination rate)을 산출하였으며, 대조구의 발아율이 90.0% 이상인 경우 실험이 적합한 것으로 판정하였다. 또한 각의 처리구에서 반응한 생물의 영향 평가는 영상처리시스템이 부착된 광학현미경(×20)으로 48시간과 72기간 동안 발아관(germ tube)의 형태가 뚜렷한 개체를 발아체로 간주하여 최소 100개체를 촬영한 후 계수하여 발아율(%, 발아한 개체수/총 개체수)를 산출하였다.
자료분석
실험 수온 구간별에서 pH 및 염분 농도별 발아율 분석은 USEPA (2002)에서 제시한 분석 과정에 따라 Toxical 5.0(Tidepool scientific Software, USA) 프로그램을 이용하여 Dunneett’s test와 maximum likelihood probit analysis 과정을 거쳐 수행하였다. 또한 각각의 수온에서 pH와 염분 노출에 따른 실험생물별 민감도는 통계처리 후 무영향농도(no observed effective concentration, NOEC), 최저영향농도(lowest observed effective concentration, LOEC) 및 반수영향농도(EC50, 50% effective concentration)를 산출한 후 비교 분석하였다.
3. 결 과
수온과 pH 농도별 실험생물의 발아율 변화
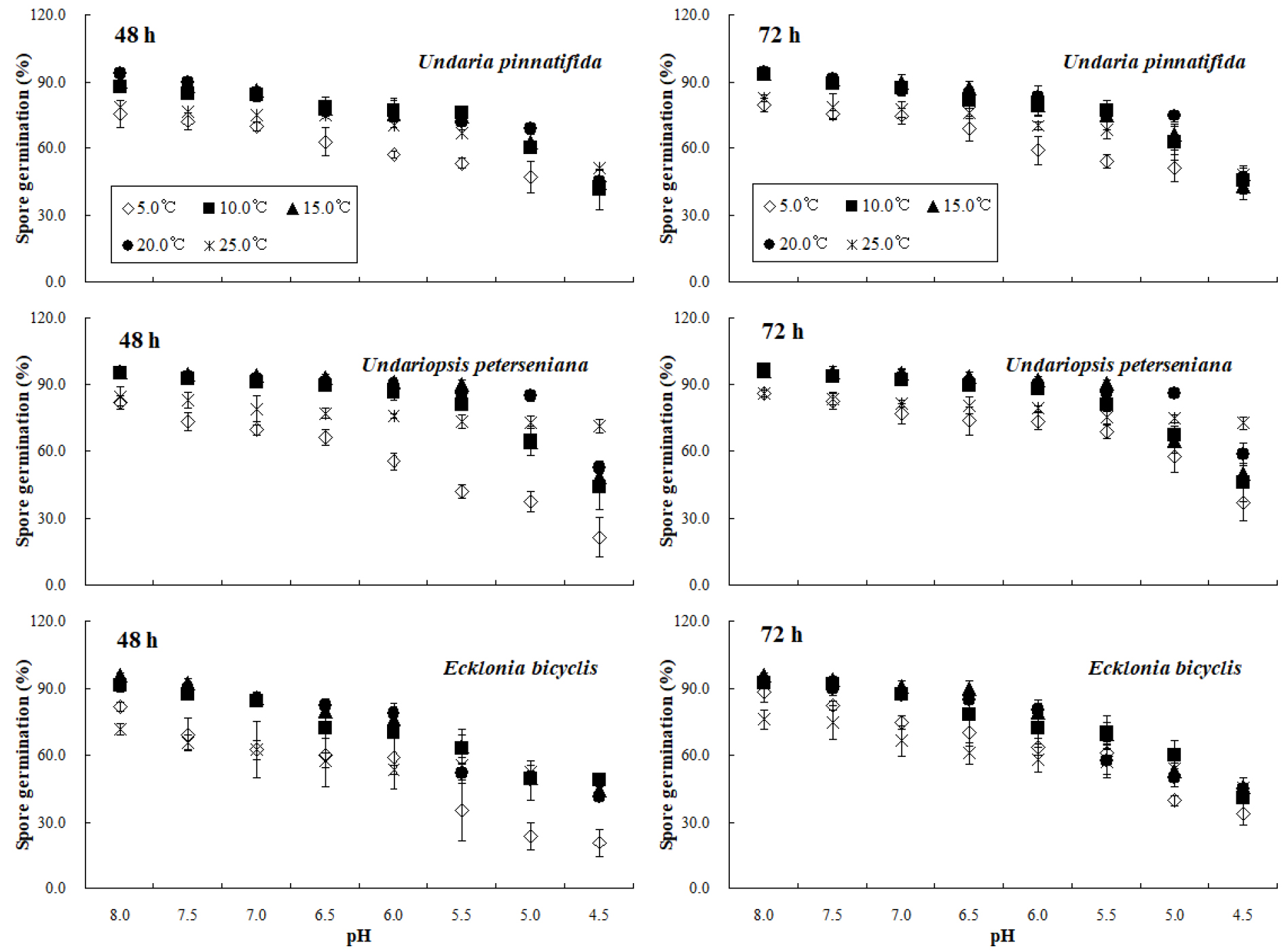
본 연구에서 pH 농도별 미역 유주자의 발아율은 5.0 °C와 25.0°C 환경에서 실험시간과 상관없이 대조군에서 90.0%를 넘지 않아 실험의 신뢰성을 확보하지 못하였다. 그러나 15.0°C와 20.0°C에서 대조군의 48시간 발아율은 각각 90.4±2.5%와 93.8±1.1%로 분석되어 신뢰성을 확보하였으며, 5.0°C와 20.0°C의 발아율은 pH 5.0보다 낮은 농도에서 50.0% 이하로 감소하였다. 넓미역의 경우 10.0°C와 15.0°C 수온에서 48시간 발아율은 pH 5.0 이하 농도에서는 50.0% 이하로 급감하였다(Fig. 2). 72시간 동안 10.0°C, 15.0°C 및 20.0°C에서 미역의 발아율은 대조군에서 90.0%보다 높게 산출되어 신뢰성을 확보하였으며, pH 5.0 보다 낮은 농도에서 50.0% 이하로 급격하게 감소하였다. 10.0°C와 15.0°C에서 넓미역의 72시간 발아율은 pH 5.5, 20.0°C에서는 pH 5.0까지 80.0%보다 높게 산출되었다. 또한 10.0°C, 15.0°C 및 20.0°C 구간의 72시간 넓미역의 발아율은 pH 4.5에서 50.0% 이하로 분석되었다. 10.0°C, 15.0°C 및 20.0°C 수온에서 대황의 72시간 발아율은 pH 6.0까지 70.0% 보다 높았으며, pH 5.5보다 낮은 농도에서는 발아한 개체가 급격하게 감소하였다. 본 연구에서 10.0°C와 15.0°C 구간에서 대황의 72시간 발아율은 pH 4.5에서 50.0% 이하로 감소하였으며, 20.0°C에서 발아율은 pH 5.0에서 50.0%보다 낮게 분석되었다(Fig. 2).
수온과 염분 농도별 실험생물의 발아율 변화
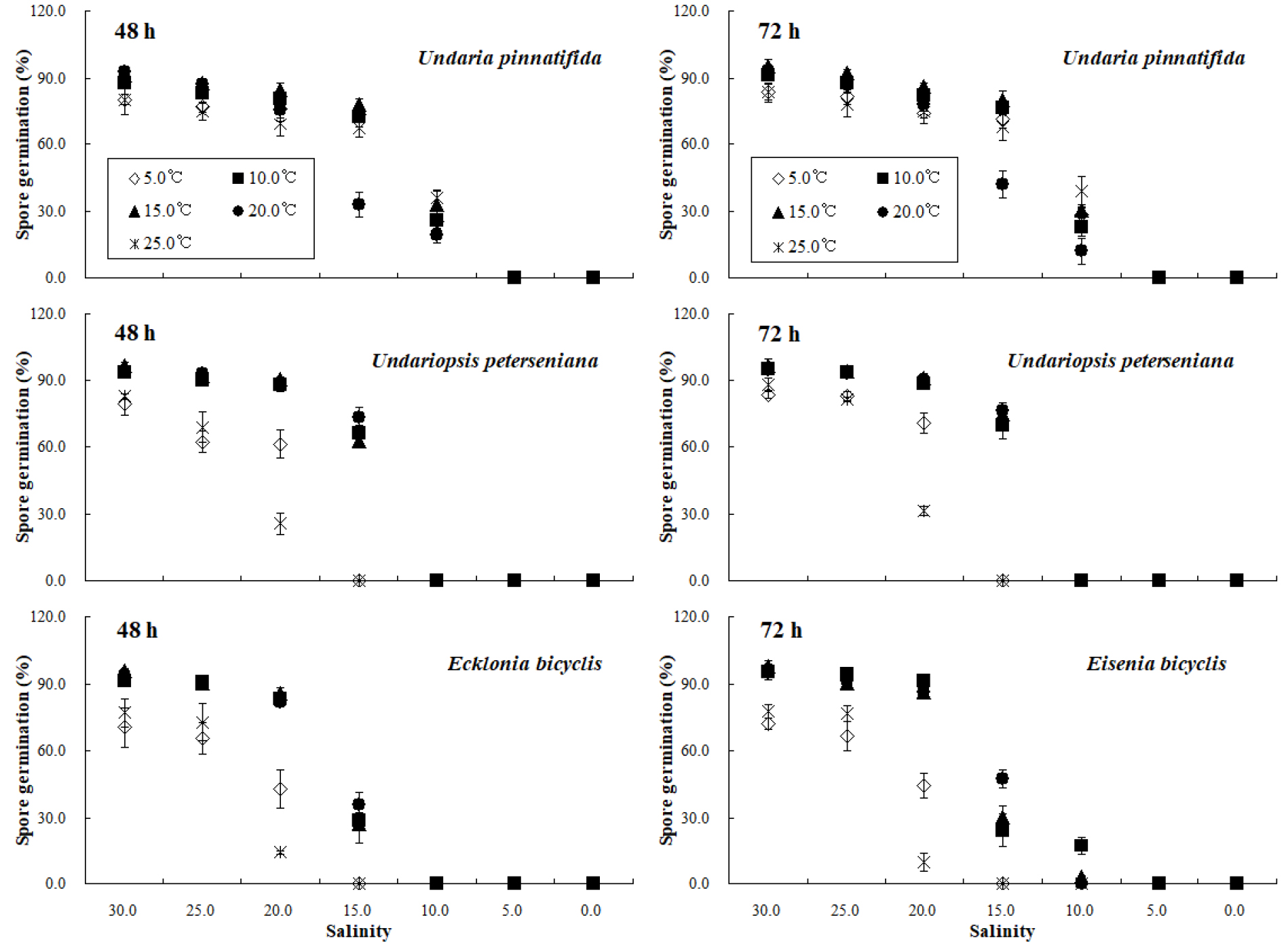
염분 농도별 실험생물의 발아율은 실험 수온에 따라 뚜렷한 차이를 보였다. 5.0°C와 25.0°C 수온에서 대형 갈조류 3종의 발아율은 pH와 유사하게 실험 시간과 상관없이 대조군에서 90.0% 이하로 산출되어 실험의 신뢰성을 확보하지 못하였다. 48시간 실험결과 10.0°C에서 미역의 발아율은 대조군에서 90.0%를 넘지 않았으나 15.0°C와 20.0°C에서는 90.0%보다 높게 분석되었다. 15.0°C와 20.0°C에서 미역 유주자의 발아율은 20.0 psu 실험구에서 80.0% 보다 높았으며, 15.0 psu 이하 농도에서는 급격하게 감소하였고, 5.0 psu에서는 수온과 상관없이 모든 개체가 사멸하였다. 10.0–20.0°C 구간에서 넓미역의 발아율은 대조군에서 90.0% 이상이었으며, 10.0 psu에서는 수온과 상관없이 모든 개체가 사멸하였다. 대황의 경우 10.0–20.0°C 구간에서 48시간 동안 15.0 psu에서 발아한 개체는 급격하게 감소하였으며, 10.0 psu에서 0.0%로 조사되었다. 또한 48시간 동안 5.0°C와 25.0°C에서 미역의 발아율은 5.0 psu, 넓미역과 대황은 15.0 psu 이하에서 0.0%가 산출되었다(Fig. 3). 72시간 염분 실험결과 10.0 °C와 15.0°C 수온에서 미역의 발아율은 10.0 psu에서 30.0% 이하로 급감하였으며, 20.0°C에서는 15.0 psu에서 42.0%의 낮은 값이 산출되었다. 넓미역 유주자는 10.0–20.0°C, 20.0 psu 조건에서 88.0%보다 많은 개체가 발아하였으며, 15.0 psu에서는 발아율 기울기가 급격히 감소하였고, 15.0 psu 이하 농도에서는 발아한 개체가 관찰되지 않았다. 대황의 경우 72시간 발아율은 20.0 psu에서 86.0%보다 높게 산출되었으며, 15.0 psu 실험구에서는 50.0% 이하로 감소하였다. 또한 10.0°C와 15.0°C에서 발아율은 10.0 psu에서 각각 17.2%와 2.8%로 산출되었으며, 20.0°C에서는 10.0 psu 이하에서는 발아한 개체가 관찰되지 않았다(Fig. 3).
수온, pH 및 염분 농도별 실험생물의 민감도 비교
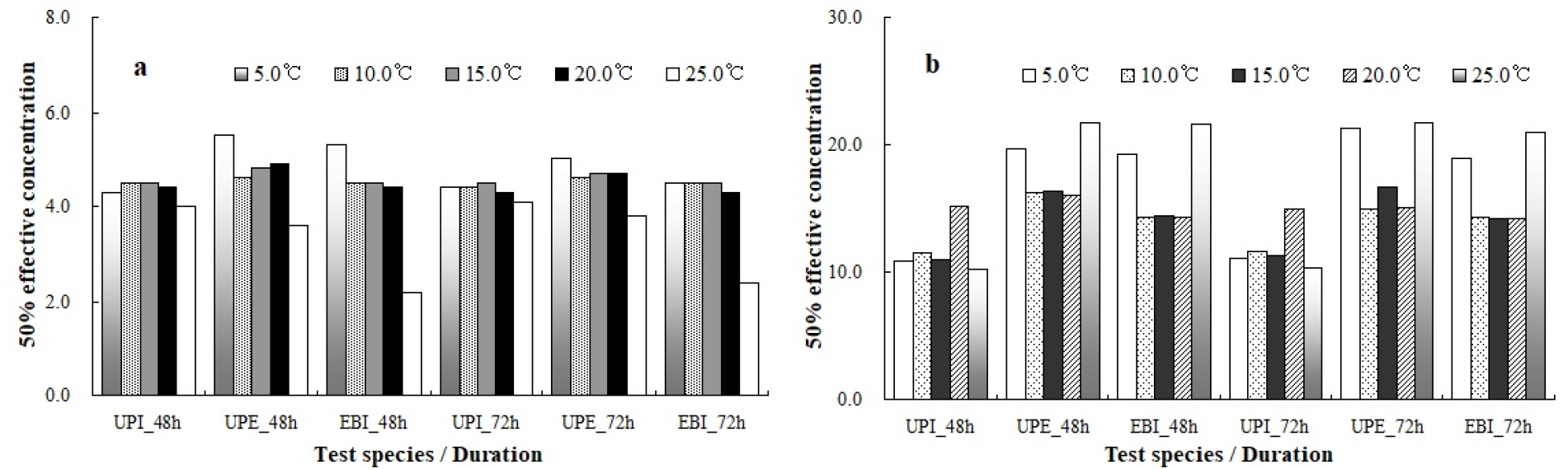
다양한 pH에서 대형 갈조류 3종의 수온별 유주자 발아율 자료를 근거로 10.0–20.0°C 구간에서 유효농도 범위를 산출한 결과, 미역의 48시간과 72시간 NOEC는 pH 7.0–7.5 범위에 존재하였으며, LOEC는 pH 6.5–7.0로 나타났다. 넓미역의 NOEC는 미역과 유사하게 pH 7.0–7.5, LOEC는 pH 6.5–7.5로 조사되었으며, 대황의 경우 48시간과 72시간 NOEC와 LOEC는 각각 pH 6.0–7.5와 pH 6.0–7.0 범위로 확인되었다. 48시간 및 72시간 EC50은 미역의 경우 10.0°C에서 각각 pH 4.5와 pH 4.4, 넓미역과 대황의 유효 농도는 실험 시간과 상관없이 각각 pH 4.6과 pH 4.5로 산출되었다. 15.0°C에서 미역의 48시간 및 72시간 EC50은 pH 4.5로 산출되었으며, 넓미역은 각각 pH 4.8와 pH 4.7로 분석되었고, 대황은 pH 4.5로 산출되어 실험 시간에 따른 농도 차이는 나타나지 않았다. pH 실험결과 20.0°C에서 미역의 48시간 및 72시간 EC50은 각각 pH 4.4와 pH 4.3으로 산출되었으며, 넓미역은 pH 4.9와 pH 4.7, 대황의 EC50은 pH 4.4와 pH 4.3으로 분석되었다(Table 1, Fig. 4).
Table 1.
Toxicological estimation of Undaria pinnatifida, Undariopsis peterseniana, and Ecklonia bicyclis exposed to pH during 48 h and 72 h at different water temperatures
| Undaria pinnatifida | Undariopsis peterseniana | Ecklonia bicyclis | |||||||
| 48 h | 72 h | 48 h | 72 h | 48 h | 72 h | ||||
| 5.0°C | NOEC* | 7.0 | 7.0 | 7.5 | < 7.5 | < 7.5 | 7.5 | ||
| LOEC** | 6.5 | 6.5 | 7.0 | 7.5 | 7.5 | 7.0 | |||
| EC50*** | 4.3 | 4.4 | 5.5 | 5.0 | 5.3 | 4.5 | |||
| 95% FL**** | 4.0–4.6 | 4.1–4.6 | 5.2–6.0 | 4.7–5.3 | 5.2–5.4 | 4.2–4.8 | |||
| 10.0°C | NOEC | 7.0 | 7.5 | < 7.5 | 7.0 | 6.5 | 7.5 | ||
| LOEC | 6.5 | 7.0 | 7.5 | 6.5 | 6.0 | 7.0 | |||
| EC50 | 4.5 | 4.4 | 4.6 | 4.6 | 4.5 | 4.5 | |||
| 95% FL | 4.4–4.6 | 4.3–4.6 | 4.1–5.0 | 4.3–4.9 | 4.4–4.6 | 4.4–4.6 | |||
| 15.0°C | NOEC | 7.0 | 7.0 | 7.5 | 7.5 | 7.0 | 6.5 | ||
| LOEC | 6.5 | 6.5 | 7.0 | 7.0 | 6.5 | 6.0 | |||
| EC50 | 4.5 | 4.5 | 4.8 | 4.7 | 4.5 | 4.5 | |||
| 95% FL | 4.3–4.6 | 4.4–4.6 | 4.6–4.9 | 4.6–4.8 | 4.3–4.7 | 4.2–4.7 | |||
| 20.0°C | NOEC | < 7.5 | 7.5 | 7.5 | 7.5 | 7.5 | 7.0 | ||
| LOEC | 7.5 | 7.0 | 7.0 | 7.0 | 7.0 | 6.5 | |||
| EC50 | 4.4 | 4.3 | 4.9 | 4.7 | 4.4 | 4.3 | |||
| 95% FL | 3.8–4.7 | 3.4–4.7 | 4.4–5.3 | 4.3–5.1 | 4.1–4.5 | 4.1–4.4 | |||
| 25.0°C | NOEC | 6.0 | 7.0 | < 7.5 | 7.0 | 7.0 | 7.5 | ||
| LOEC | 5.5 | 6.5 | 7.5 | 6.5 | 6.5 | 7.0 | |||
| EC50 | 4.0 | 4.1 | 3.6 | 3.8 | 2.2 | 2.4 | |||
| 95% FL | 3.6–4.2 | 3.8–4.3 | 2.9–4.1 | 3.2–4.2 | 0.8–3.0 | 1.0–3.1 | |||
염분 농도별 미역의 48시간 및 72시간 NOEC는 10.0°C에서 25.0 psu로 산출되었으며, 넓미역은 48시간 동안 25.0 psu, 72시간 NOEC는 20.0 psu로 조사되었고, 대황의 NOEC는 48시간과 72시간 동안 25.0 psu로 분석되었다. 한편 15.0°C와 20.0°C 환경에서 실험생물 3종의 NOEC는 실험 시간과 관계없이 25.0 psu 부근에 존재하는 것으로 조사되었다. LOEC의 경우 10.0°C 수온 구간에서 미역은 실험 시간과 상관없이 20.0 psu로 산출되었으며, 넓미역은 25.0 psu, 대황의 경우 48시간 동안 25.0 psu, 72시간 실험에서는 20.0 psu로 분석되었다. 20.0°C에서 미역의 48시간과 72시간 LOEC는 각각 25.0 psu로 산출되었으며, 넓미역은 48시간 동안 20.0ps, 72시간 LOEC는 25.0 psu로 조사되었으며, 대황의 LOEC는 실험 시간과 상관없이 20.0 psu 부근에 존재하였다. 실험생물의 EC50을 산출한 결과 10.0°C에서 미역의 48시간 EC50은 11.5 psu, 72시간 EC50은 11.6 psu로 조사되었으며, 넓미역은 각각 16.2 psu와 15.0 psu로 분석되었고 대황은 실험 시간과 상관없이 14.3 psu로 산출되었다. 15.0°C에서 미역의 48시간 EC50은 11.0 psu, 72시간에는 11.3 psu로 조사되었으며, 넓미역은 각각 16.3 psu와 16.7 psu, 대황은 14.4 psu와 14.2 psu로 분석되었다. 20.0°C에서 미역의 48시간 EC50은 15.2 psu로 조사되었으며, 72시간 실험결과 15.0 psu로 산출되었다. 넓미역의 48시간과 72시간 EC50은 각각 16.0 psu와 15.1 psu, 대황의 EC50은 14.3 psu와 14.2 psu로 분석되었다(Table 2, Fig. 4).
Table 2.
Toxicological estimation of Undaria pinnatifida, Undariopsis peterseniana, and Ecklonia bicyclis exposed to salinity during 48 h and 72 h at different water temperatures
| Undaria pinnatifida | Undariopsis peterseniana | Ecklonia bicyclis | |||||||
| 48 h | 72 h | 48 h | 72 h | 48 h | 72 h | ||||
| 5.0°C | NOEC* | 15.0 | 25.0 | 25.0 | < 25.0 | < 25.0 | 25.0 | ||
| LOEC** | 10.0 | 20.0 | 20.0 | 25.0 | 25.0 | 20.0 | |||
| EC50*** | 10.9 | 11.1 | 19.6 | 21.2 | 19.2 | 18.9 | |||
| 95% FL**** | 10.6-11.4 | 10.7-11.5 | - | 20.7-21.8 | - | 9.5-19.5 | |||
| 10.0°C | NOEC | 25.0 | 25.0 | 25.0 | 20.0 | 25.0 | 25.0 | ||
| LOEC | 20.0 | 20.0 | 20.0 | 15.0 | 20.0 | 20.0 | |||
| EC50 | 11.5 | 11.6 | 16.2 | 15.0 | 14.3 | 14.3 | |||
| 95% FL | 11.0-11.9 | 10.6-13.1 | 15.8-16.5 | - | - | - | |||
| 15.0°C | NOEC | 25.0 | 25.0 | < 25.0 | < 25.0 | < 25.0 | 25.0 | ||
| LOEC | 20.0 | 20.0 | 25.0 | 25.0 | 25.0 | 20.0 | |||
| EC50 | 11.0 | 11.3 | 16.3 | 16.7 | 14.4 | 14.2 | |||
| 95% FL | 10.6-11.4 | 10.3-12.5 | 16.0-16.6 | 16.3-17.1 | 10.4-14.7 | - | |||
| 20.0°C | NOEC | < 25.0 | < 25.0 | 25.0 | < 25.0 | 25.0 | 25.0 | ||
| LOEC | 25.0 | 25.0 | 20.0 | 25.0 | 20.0 | 20.0 | |||
| EC50 | 15.2 | 15.0 | 16.0 | 15.1 | 14.3 | 14.2 | |||
| 95% FL | 11.8-20.7 | 14.6-15.5 | 15.6-16.3 | 14.8-15.4 | - | - | |||
| 25.0°C | NOEC | 25.0 | 25.0 | 25.0 | 25.0 | < 25.0 | < 25.0 | ||
| LOEC | 20.0 | 20.0 | 20.0 | 20.0 | 25.0 | 25.0 | |||
| EC50 | 10.3 | 10.4 | 21.7 | 21.7 | 21.6 | 20.9 | |||
| 95% FL | 9.9-10.8 | 9.9-10.9 | 21.2-22.4 | 21.1-23.4 | 21.1-22.1 | 20.5-21.3 | |||
4. 고 찰
해수의 수온 상승은 생물의 생리적, 생화학적 대사과정에 영향을 주어 종의 분포 및 다양성뿐만 아니라 생태계 기능에 영향을 미치는 것으로 보고되었다(Oliver et al. 2018; Darmaraki et al. 2019; Smale et al. 2019; Garrabou et al. 2022). 수온과 더불어 해수의 화학적 특성 변화 및 담수 유입으로 인해 야기되는 해수의 pH 및 염분 변화는 해조류의 성장과 생리 활성에 영향을 줄 수 있음에도 불구하고 2종 이상의 환경인자가 조합된 영향 평가는 제대로 이루어지지 않았다(Choi et al. 2019; Yoon 2023). 이에 본 연구에서는 해양환경 변화에 따른 해조류의 생태-생리 변화를 예측하고자 미역, 넓미역 및 대황 등 대형 갈조류 3종의 초기 생활사에 미치는 수온 구간별 pH 및 염분 영향을 평가하였다.
본 연구에서 5.0°C와 25.0°C에서 대조군의 발아율은 미역, 넓미역 및 대황 모두 90.0%를 넘지 않았으며, 30.0 °C에서는 실험 물질 및 시간과 상관없이 모든 개체가 사멸하였다. 일반적으로 국내뿐만 아니라 유럽 및 미국에서는 생물검정 시 생물의 적합성 유지를 위한 시험적합도기준(test acceptability criterion)을 설정하고 있으며, 생물의 사망 또는 사멸률 시험의 경우 대조군에서 실험 완료시까지 시험생물의 90.0% 또는 그 이상 생존하는 조건을 제시하고 있다(ASTM 1996; NIWA 1998; USEPA 2002; MOF 2018). 이에 본 연구에서 5.0°C, 25.0°C 및 30.0°C 수온에서 수행된 실험은 대조군의 생존률이 90.0 %를 넘지 못하였으므로 시험적합도 기준을 충족하지 못하였다. 한편 5.0°C와 25.0°C에서 실험생물의 발아율은 실험 물질과 상관없이 수온에 따라 차이를 보였는데, 5.0 °C에서는 사멸하는 개체 없이 발아율이 낮았으며, 25.0 °C에서는 대부분의 유주자가 사멸하였다. 본 연구에 사용된 3종의 대형 갈조류와 다시마(Saccharina japonica)는 전형적인 이형 생활사를 가지는 종으로써 10.0–15.0°C의 낮은 수온에서 주로 성장하고 25.0°C보다 높은 수온에서는 생존할 수 없으며, 고수온은 대사 활동에 영향을 받아 포자와 배우체의 사멸을 초래할 뿐만 아니라 생존한 포자와 배우체의 성장을 억제하는 것으로 보고된 바 있다(Choi et al. 2019). 이와 같은 현상은 본 연구에서도 유사하게 관찰된 바 있는데, 대형 갈조류 3종의 경우 수온 스트레스에 민감한 반응을 보이는 것으로 판단되었다. 특히 포자와 배우체는 고수온에 대한 내성이 급격하게 감소하여 결국 사멸하는 개체가 증가하는 것으로 보고되었는데(Zhang et al. 2022a), 본 연구에서 5.0°C의 저수온 환경에서는 실험생물의 생리 활성도가 낮아 적정 수온에 도달하기까지 발아가 지연된 것으로 판단되며, 25.0°C의 고수온 환경에서는 내성 한계를 벋어남에 따라 해조류가 녹기 시작하는 단계이므로 발아체 및 배우체가 형성되지 않는 것으로 해석할 수 있다. 한편 열대지역에 서식하는 해조류는 고수온보다 저수온에 대한 영향을 받게 되는데 저온 스트레스는 세포막 구조를 손상시키고 지질 구성을 변화시키며 광합성 능력을 약화시키는 것으로 보고되었다(Matteucci et al. 2011; Theocharis et al. 2012). 또한 뉴질랜드의 고유종인 Lessonia variegata는 고수온에 대한 민감도가 높아 25.0°C에서 광합성 수율과 chloroplyll-a의 농도 감소와 같은 생리 작용에 부정적인 영향을 보였다(Bollen et al. 2016). 이와 같은 현상은 대형 갈조류 3종의 경우 수온에 대한 내성 범위를 벋어난 경우 단기간 동안 고수온 환경에서 생리 상태의 변화만 초래하지만 장기간 동안 고수온 스트레스에 환경에서는 결국 사멸하는 것이 공통적인 현상으로 판단되었다.
본 연구에서 대형 갈조류 3종을 대상으로 10.0–20.0°C 수온 구간에서 생물의 발아율은 실험 물질의 농도-반응 사이의 기울기 변화가 뚜렷하였다. 발아율은 실험 시간에 따라 다소 차이는 있었으나 10.0–20.0°C 구간에서 72시간 실험한 미역의 발아율은 pH 5.0 보다 낮은 농도에서 50.0% 이하로 급격하게 감소하였으며, 넓미역은 pH 4.5에서 50.0% 이하로 분석되었다. 한편 대황의 경우 발아율은 10.0°C와 15.0°C 구간에서 pH 4.5, 20.0°C 환경에서는 pH 5.0로 산출되었는데, 본 연구결과 대형 갈조류 3종의 발아율은 pH 4.5~5.0 이하 농도에서 50.0% 이하로 발아율이 급격히 감소하는 것으로 분석되었다. 해조류의 성장과 대사 활동은 주로 CO2를 이용한 광합성을 통해 이루어지는데, 해수의 pH와 밀접한 관계가 있는 CO2 농도 변화는 해양산성화 뿐만 아니라 해조류의 초기 성장과 광합성에 긍정적 및 부정적 영향을 줄 수 있는 것으로 보고되었다(Xiao et al. 2021). 특히 pH 변동은 석회화 생물에게 적응 기회를 제공하며, 해양산성화에 따른 산호류의 반응 연구에 도움을 줄 수 있기 때문에 pH가 낮거나 pH 변동이 더 잦은 미래 해양환경에 대한 생태계 변화를 예측할 수 있는 잠재력이 높은 것으로 보고되었다(Ramajo et al. 2019). 한편 본 연구에서 10.0–20.0°C 수온 구간에서 pH 농도별 대형 갈조류 3종의 유주자 발아율에 대한 유효농도 범위를 산출하였다. 분석 결과 10.0°C에서 72시간 EC50은 pH 4.4–4.6, 15.0°C 수온 구간에서는 pH 4.5–4.7, 20.0°C에서는 pH 4.3–4.7로 산출되었다. 또한 발아에 유효한 영향을 미치는 72시간 EC50은 넓미역이 가장 높았으며, 미역과 대황은 큰 차이를 보이지 않아 수온과 상관없이 넓미역이 미역과 대황보다 pH 변화에 대한 민감도가 상대적으로 높은 것으로 분석되었다. 해수의 수온 상승과 pH 변화는 해양생물의 생태-생리에 큰 영향을 미치는 것으로 알려져 있다(Arbit et al. 2018; Stipcich et al. 2023). 특히 수온 상승과 낮은 pH 농도가 동시에 발생하는 환경은 해양생물의 성장과 생존율에 심각한 영향을 미치는 것으로 알려져 있으며, 특정 종의 감소와 생물학적 상호작용의 변화와 같이 군집 구성에 직간접적인 영향 미치는 것으로 예측되었다(Wernberg et al. 2016). 고수온 환경에서 낮은 pH와 강한 조도는 다시마의 배우체에 저항성을 감소시켜 생식활동에 부정적인 영향을 미치며, 장기적으로는 해조류 군집 구조가 바뀔 가능성이 제시된 바 있다(Xu et al. 2015). 본 연구에서 설정한 실험조건에 다소 차이가 있으나 giant kelp Macrocystis pyrifera는 수온 18.0°C, pH 7.46 환경조건에서 발아율을 크게 감소시키고, 유주자의 사망률을 증가시키는 것으로 보고된 바 있다(Gaitán-Espitia et al. 2014). 이와 같은 현상은 이형 생활사를 거치는 해조류에서 발생 가능한 것으로 제시되었는데, 본 연구에서 실험생물로 사용된 미역, 넓미역 및 대황은 다시마와 동일한 이형 생활사 단계를 거치는 종으로써 고수온과 낮은 pH 환경은 수온 및 pH 변화에 따른 스트레스로 인해 생물의 발아, 성장 및 대사 활동과 같은 다양한 생태-생리 변화를 유발할 것으로 판단된다.
다양한 수온 환경에서 염분 농도와 발아율은 뚜렷하게 구분되었는데, 10.0–20.0°C 수온에서 72시간 동안 미역의 발아율은 10.0–15.0 psu, 넓미역은 15.0 psu에서 50.0 % 이하로 기울기가 급격히 감소하였고 15.0 psu 이하 농도에서는 발아한 개체가 관찰되지 않았다. 또한 대황의 발아율은 15.0 psu에서 50.0% 이하로 감소하였으며 10.0 psu 보다 낮은 농도에서는 발아한 개체가 관찰되지 않았다. 해양에서 염분 감소는 해조류의 성장, 광합성, 세대 기간 및 생식에 영향을 미치는 것으로 보고되었다(Gaitán- Espitia et al. 2014; Bews et al. 2021). 예를 들면, 다시마는 염분이 감소하면 저염분 스트레스로 인해 성장 속도와 색소 감소가 유발되는 것으로 제시되었는데, 34.0 psu에서 10.0 psu로 염분이 하강하면 광합성 효율이 뚜렷하게 감소하였다(Kroeker et al. 2010; Raybaud et al. 2013). 본 연구에서 수온 구간별 갈조류 3종의 염분에 대한 민감도를 비교한 결과 미역의 경우 72시간 EC50은 10.0°C에서 11.6 psu로 조사되었으며, 넓미역과 대황은 각각 15.0 psu와 14.3 psu로 산출되었다. 15.0°C 구간에서는 미역이 11.3 psu, 넓미역은 16.7 psu, 대황은 14.2 psu로 분석되었으며, 20.0°C에서 미역의 72시간 EC50은 15.0 psu, 넓미역은 15.1 psu, 대황의 경우 EC50은 14.2 psu로 분석되었다. 한편 수심이 낮은 지역에 서식하는 해조류는 저염 및 고염분 스트레스에 민감한 것으로 보고되었는데(Bollen et al. 2016), 20.0–40.0 psu 사이에서 협염성 광합성 반응을 수행하는 다시마는 저염분 환경에서 광합성 활동이 감소하였다. 이러한 현상은 극심한 저염 또는 고염분 스트레스 하에서 세포질은 이온 농도를 증가시키고 선택적 흡수로 인해 이온 비율도 변화하며, 이러한 스트레스 하에서 광합성과 호흡이 억제될 수 있는 것으로 보고되었다(Karsten 2012). 본 연구에서 대형 갈조류 3종을 대상을 염분에 대한 민감도를 비교한 결과 대부분의 생물은 10.0–15.0 psu 범위에서 저염분 스트레스가 시작되는 것으로 판단되었으며, 10.0–20.0°C 구간에서 생물의 발아에 영향을 미치는 50% 유효농도는 수온과 상관없이 넓미역이 염분 변화에 가장 민감하게 반응하였으며 다음으로 대황은 10.0°C와 15.0°C, 20.0°C에서는 미역의 민감도가 대황보다 높은 것으로 분석되었다. 이와 같은 결과는 저염분 환경에서 세포 내 이온 교환이 교란됨으로써 광합성 및 영양염 흡수를 통한 생물의 생식, 성장 및 분포 등의 생리-생태 활성에 부정적인 영향을 미치는 것으로 해석되었다(Zhang et al., 2022b). 따라서 본 연구에 사용된 대형 갈조류 3종의 발아율은 10.0–15.0 psu 구간에서 급격하게 감소하여 저염분 스트레스에 대한 생물의 생태-생리 반응은 유사하게 관찰되었는데, 이는 삼투 스트레스에 따른 세포 간 Na+, K+, Cl- 등의 이온 농도 변화와 연관된 것으로 판단되었다(Zhang et al. 2008). 한편 수온과 염분 등의 여러 가지 결합된 스트레스 요인의 효과는 대개 단일 작용의 단순한 가산 효과, 즉 “교차 시너지즘”을 유발할 수 있는데(Alexieva et al. 2003), 예를 들면, 미역의 유주자는 8.0 psu에서 발아하고, 배우체는 6.0 psu의 낮은 농도에서 생존 가능한 반면, 어린 엽체(최대 길이 4 mm)는 16.0 psu까지 생존 가능하며, 성숙한 포자체는 5.0–20.0°C 수온 범위에서 10일 동안 6.0 psu의 저염분 조건에서도 생존이 가능한 것으로 보고되었다(Bollen et al. 2016). 이와 같은 결과는 실험환경이 다소 상이한 본 연구결과와 다소 차이를 보였으나 적절한 수온과 염분의 조합은 해조류 생활사 단계별로 생리적 내성을 유지할 수 있는 잠재적인 상승효과로 작용할 수 있음을 시사한다.
대형 갈조류로 분류되는 해조류 중 미역의 암배우체 생존율과 상대성장률의 최적 조건은 수온 15.0°C, 염분 30.0 psu, pH 8.0, 광도 60 µmol photon m-2s-1로 분석되었으며, 대황의 경우 최적 성숙 온도는 15.0–20.0°C 범위이며, 20.0°C에서 가장 생장이 높은 것으로 보고되었다(Lee and Kim 2020; Lee et al., 2022). 또한 넓미역 암수배우체는 생장 후 성숙되기 때문에 10.0–15.0°C, 10–20 µmol photon m-2s-1에서 촉진되는 것으로 분석되었다(Hwang et al. 2010a). 이와 같은 결과는 이형생활사를 가진 갈조류의 아포체 형성율을 높이기 위한 생존전략으로써 성장단계별로 유리한 수온과 광 조건에서 생물의 활성 변화를 인식하는 메커니즘이 존재할 것으로 제시되었다(Hwang et al., 2011). 본 연구에서 10.0–20.0°C의 수온 구간에서 미역, 대황 및 넓미역의 발아율은 pH 4.5–5.0, 10.0–15.0 psu에서 급격히 감소하였으며, 5.0°C와 25.0°C에서는 발아가 지연되거나 발아한 개체는 거의 관찰되지 않았고, 30.0°C에서는 모든 개체가 사멸하였다. 또한 EC50은 pH 4.4–4.7과 11.6–16.7 psu로 조사되었으며, 염분과 pH에 대한 실험생물의 민감도는 수온과 상관없이 넓미역이 가장 높았고, 미역과 대황은 큰 차이를 보이지 않았다. 이와 같은 민감도의 차이는 실험 종의 서식환경에 따라 차이가 나는 것으로 판단되는데, 미역과 대황의 경우 조하대 5 m 수심 이내에 분포하며 다양한 환경변화에 노출되는 빈도 또는 강도가 높아 이에 저항할 수 있는 대사 능력이 발달한 것으로 해석된다. 이와 반면 20 m 이하 수심에 서식하는 넓미역은 미역이나 대황보다 서식환경이 안정된 깊은 수심에서 대사 활동을 유지하기 때문에 낮은 pH와 염분에 대한 적응능력이 낮아 상대적으로 환경변화에 대한 민감도가 높은 것으로 판단할 수 있다. 결론적으로 pH나 염분에 노출된 미역, 대황 및 넓미역의 환경 스트레스 및 내성은 해수의 수온, pH 및 염분의 조합된 변화에 따라 잠재적으로 상승 또는 하강할 수 있으며, 이와 같은 효과는 해조류의 종 및 생활사 단계에 따라 차이를 보이는 것으로 판단된다. 또한 환경스트레스에 노출된 생물의 민감도는 생물 종, 서식 수심, 해수의 증발량, 연안 지역의 담수 유입 강도에 따라 직, 간접적으로 차이를 보이는 것으로 예측할 수 있다. 더불어 본 연구에서 서식환경이 상이한 대형 갈조류 3종을 대상으로 다양한 수온 구간에서 수행한 해양생태독성평가 결과는 기후변화, 해양산성화 및 갯녹음 발생에 따른 해조류의 분포 및 생리-생태 변화를 예측할 수 있는 자료로써 활용가치가 높은 것으로 판단된다.
5. 적 요
본 연구에서는 대형 갈조류인 미역(Undaria pinnatifida), 넓미역(Undariopsis peterseniana)과 대황(Ecklonia bicyclis)의 초기 생활사에 대한 수온, pH 및 염분이 조합된 영향을 평가하기 위해 해양생태독성평가를 수행하였다. 실험결과, 10.0–20.0°C 구간에서 pH와 염분 농도별 유주자의 발아율은 실험 물질의 농도-반응 사이에 선형관계가 뚜렷하였다. 실험기간 동안 미역, 넓미역 및 대황의 발아율은 pH 4.5–5.0과 10.0–15.0 psu에서 급격히 감소하였다. 또한 72시간 EC50은 pH 4.4–4.7, 11.6–16.7 psu로 조사되었으며, 실험 물질에 대한 민감도는 수온과 관계없이 넓미역이 가장 높았고, 미역과 대황은 유의한 차이를 보이지 않았다. 결론적으로 pH나 염분에 대한 미역, 대황 및 넓미역의 환경 스트레스 및 내성은 해수의 수온, pH 및 염분의 조합된 변화에 따라 잠재적으로 상승 또는 하강할 수 있으며, 해조류의 종 및 생활사 단계에 따라 차이를 보이는 것으로 해석된다. 본 연구결과는 기후변화, 해양산성화 및 갯녹음으로 인한 해초의 생태-생리적 변화를 예측하기 위한 자료로서 활용가치가 높은 것으로 판단된다.
